11.2. Почему «процветают» примитивные паразиты
11.2. Почему «процветают» примитивные паразиты
Одним из главных материалистических достижений дарвинизма явился отказ от признания ламарковского внутреннего стремления к совершенству как одной из причин эволюции. (В гл. 1 мы говорили об отношении Ч. Дарвина к прогрессу, который считал, что нет направленного прогресса, нет повышения организации в эволюции.) Однако в целом повышение организации живых структур в макроэволюции явно прослеживается, о чем говорит развитие жизни от пробионта до высшего животного или растения. Отсюда и возникает неудовлетворенность дарвинизмом, поскольку повышение приспособленности, как правило, связано с усложнением структур, а в дарвинизме этого не объясняется. В то же время хорошо известны и широко распространены случаи дегенерации форм, потери ими тех сложных надстроек, которые приобретались в процессе длительного действия отбора на выживание наиболее приспособленных. Самоорганизация жизни, лежащая в основе представлений дарвинизма о движущих силах развития, никак не может объяснить явление дегенерации форм организмов в эволюции. (Зачем нужно упрощение на общем пути к самоусложнению?) Отсюда—прямой путь к агностицизму и к признанию непредсказуемости действия естественного отбора. Получается, что отбор делает, что хочет («выступает в роли господа бога» — по А. А. Любищеву), то усложняя, то, наоборот, резко упрощая структуру. Остается только post factum фиксировать все то, что он «напридумывал» в результате «мгновенного принятия решения», т. е. перейти на описательный путь и попытаться классифицировать имеющееся. Это и было тщательно проделано дарвинистами при разработке учения о разных формах прогресса. Наиболее обстоятельно учение о прогрессе освещено в трудах А. Н. Северцова [1922,1967].
Он был вынужден признавать, что общая дегенерация является полной противоположностью морфофизиологического прогресса, или ароморфоза. Действительно, у многих дегенерировавших форм целиком редуцируются органы движения, т. е. вся мускульная система, а также скелет конечностей, редуцируется центральная нервная система со всеми нервами и органами чувств и органы активного питания и, наоборот, прогрессивно развивается половая система [Северцов, 1967]. Очень существенным замечанием А. Н. Северцова является то, что он говорит о понижении и самой «интенсивности проявления жизнедеятельности при дегенерации вместе с понижением сложности организации» [Там же, т. 138]. Таким образом, дегенерация и ароморфоз полностью противопоставляются по направлению изменений как структуры, так и функции. Но тем не менее оба типа изменений могут приводить к процветанию вида, а это и означает, что их следует включить в эволюционный процесс как равноценные направления, приводящие к биологическому прогрессу (вместе с идиоадаптациями и ценогенезами). Что же, на этом пути остается еще более абстрагироваться, признать единство противоположностей и перестать искать естественно-научное, в данном случае биологическое, объяснение различия форм прогрессивного развития. По-видимому, в пределах только субстратного подхода не удается сделать большего, чем констатация имеющегося. Об этом говорил и А. Н. Северцов [1967], ограничивая рассмотрение эволюции животных тем, «как она происходит в действительности» и не рассматривая, «почему происходит именно то, а не другое изменение» (с. 125).
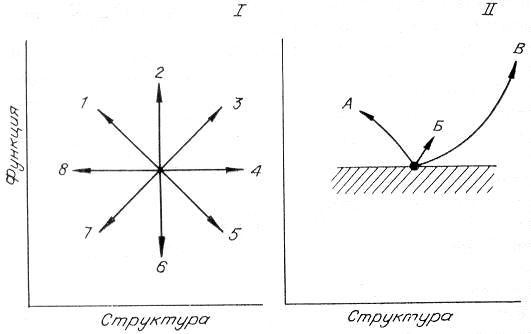
Рис. 18. Схема возможных изменений в системе «структура — функция» организма (I) и разрешенные (с позиций С + Э подхода) изменения структуры и функции в эволюции: А — направление дегенерации, Б — идиоадаптации и ценогенезы, В — ароморфозы (II). Заштрихована запрещенная зона.
Попробуем, в дополнение к субстратному, применить энергетический подход, т. е. использовать С + Э концепцию. Само собой разумеется, что всякое эволюционное изменение — это изменение и структуры, и функции организма. Схематически все возможные сочетания таких изменений представим в фазовой плоскости «структура—функция» (рис. 18). Стрелка 3 означает, что усложнение структуры связано с усложнением функции, в нашем случае — с увеличением потока энергии, перерабатываемого данным типом биологической структуры. Это как раз и соответствует прогрессивному изменению с позиций структурного подхода. Направлениями от стрелки 2 до стрелки 4 с разными углами наклона и будут иллюстрироваться прежде всего ароморфозы. Заметим, что сюда же следует отнести и идиоадаптации, так как энергетический подход требует хоть и небольшого, но все же усиления функции. Начиная со стрелки 4, включая 5, 6, 7 и 8, мы имеем явное ухудшение функции, что запрещается энергетическими принципами и потому не должно реализовываться. «Самыми запрещенными» являются направления, близкие к стрелке 5, т. е. усложнение структуры и ослабление функции.
Направления, примыкающие к стрелке 1, и будут характеризовать явление дегенерации, так как структура явно упрощается. Но при этом энергетический подход требует усиления функции и не разрешает ее ослабления (по крайней мере в удельном выражении). Термин «общее понижение жизнедеятельности» требует здесь некоторого уточнения. Если организм, популяция вынуждены переходить в новые, часто более простые условия существования (пещеры, большая глубина воды, отсутствие света, глубокие илы и т. д.), то по сравнению со старым местообитанием его функционирование ухудшается. И по-видимому, только упрощение структуры, избавление от ненужных на новом месте старых приспособлений позволяет сохранить и одновременно интенсифицировать функцию, т. е. повысить жизнедеятельность. Таким образом, учет энергетического подхода резко сужает разрешенные варианты развития в системе «структура — функция». Кстати, снимаются и противоположные направления, т. е. тот произвол, что был в рамках структурного подхода (см. рис. 18, II).
Очень существенно то обстоятельство, что эволюция «вниз» по функции не разрешается, если учитывать энергетические принципы. Поэтому говорить о том, что «микроорганизмы эволюционируют в стороны и вниз, но не вверх», с позиций С + Э подхода некорректно, ибо функция улучшается — такая трактовка допустима только с позиций субстратного подхода. Современные микроорганизмы отличаются от древних форм не только разнообразием структур (и иногда их серьезным упрощением), но и существенным усложнением функций: сюда входят и использование различных источников энергии и вещества, недоступных высшим формам, и интенсификация функции до уровней, тоже недостижимых для высших организмов. Но, совершенствуя и во многих случаях упрощая структуру, они под действием естественного отбора интенсифицируют функцию, т. е. эволюционируют «вверх». Как мы обсуждали в предыдущем разделе, питание растворенными веществами делает невыгодным увеличение размеров именно с позиций энергетики. Подчиняясь этому, прокариотные формы выполняют свою миссию в круговороте, «не беспокоясь» об усложнении структур, а наоборот, в соответствии с энергетическими принципами, «заботясь» об освобождении от любой структурной избыточности. Именно на микроорганизмах может быть ярче всего продемонстрирована способность необычайно быстро избавляться от лишних структур.
Как замечает Э. Брода в книге «Эволюция биоэнергетических процессов» [М., 1978, с. 61–62]: «Организмы имеют тенденцию утрачивать ненужные гены и ферменты. Это явление играет огромную роль в эволюции, и особенно в эволюции микробов...
...Автотрофы имеют тенденцию превращаться обратно в гетеротрофы, если дать им подходящие питательные вещества. И вообще, готовность к утратам — это свойство клеток, которое было хорошо изучено на простых системах». Одним из наиболее изученных и показательных фактов «гибкости» приспособления на уровне микробных популяций является смена прототрофных форм в популяции на ауксотрофные. Обычно на обедненных средах прототрофные варианты, способные производить для себя все необходимые метаболиты, быстро вытесняют ауксотрофные формы, требующие для своего развития дополнительно какого-либо определенного соединения. Если же это соединение добавлено в среду, то ауксотроф получает преимущество и способен расти быстрее, так как более экономен и не производит этого соединения в отличие от прототрофов. Этот пример с победой «дефективного» ауксотрофа при развитии на обогащенной среде может служить хорошей моделью явления дегенерации формы при переходе к паразитизму. (Дефективность, т. е. упрощение структуры ауксотрофа, может быть связана с потерей гена (делецией) в ДНК клетки, т. е. иметь генетическую природу.)
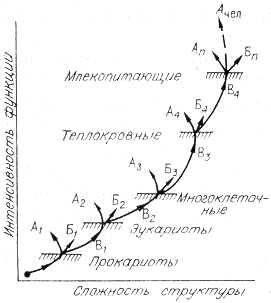
Рис. 19. Общая схема относительного изменения структуры и функции организмов в макроэволюции.
1 — прокариоты; 2 — эукариоты; 3 — многоклеточные; 4 — теплокровные; n — млекопитающие. Буквенные обозначения те же, что и на рис. 18.
Покажем схему усложнения структуры и функции организмов, начиная с пробионтной клетки и кончая млекопитающими, в том числе и человеком (рис. 19). В этом ряду несомненно усложнялась структура, но закреплялись только те варианты, которые приводили к существенному усилению функции, как того требует С + Э подход. Практически на каждом этапе, связанном с крупным ароморфозом, помимо усложнения структуры наблюдалась и дегенерация формы (структуры). На рисунке это отмечено стрелками А, загнутыми влево и вверх. Заметим, что к стрелкам А, указывающим на дегенерацию структуры млекопитающих, по-видимому, должна примыкать и стрелка Ачел, соответствующая человеку. Действительно, что касается структуры, то мы в эволюции немало потеряли: шерсть, спасающую от холода; острые клыки хищника; возможно, и хвосты, помогающие предкам лазать по деревьям. Но энергетика человека как вида возросла в десятки раз (см. гл. 9).
Общая тенденция увеличения угла наклона кривой отражает тот факт, что с повышением организации функция растет быстрее, и это во многом связано с цефализацией структуры. Следовательно, для более точных представлений о темпах эволюции необходимо привлекать и информационный подход. К этому же можно добавить, что возможность какого-либо ароморфоза у млекопитающих, связанного с сильным изменением структуры, мало вероятна. Эволюция из биологической трансформировалась в технологическую, особенно если иметь в виду энергетику человечества. Став глобальной силой, человечество способно вмешиваться в ход эволюции, переведя ее на направленный отбор, селекцию и регулируемую перестройку генотипов.
Хотелось бы обратить внимание на всеобщность и распространенность явления конвергенции, во многих случаях тесно связанного с явлением регресса формы. Конечно же, общими физическими законами определяются особенности и конкретные проявления конвергентности признаков на различных уровнях организации. Среди этих законов хорошо прослеживается действие обоих энергетических принципов. Как правило, конвергентное развитие приводит к наилучшему структурному варианту, позволяющему организму как можно больше изымать энергии (пищи) из окружения, при этом минимизируя собственные траты энергии на перемещение и ориентацию в пространстве и его использование.
Именно сходство условий существования определяет сходство форм и структур различных, а иногда и очень отдаленных таксонов. Это значит, что сходные признаки контролируются очень разными генами или генными комплексами. А это, в свою очередь, значит, что концепция: «Гены хотят жить и размножаться» — явно не состоятельна. Структура оказывается вторичной, подчиняясь функции, которая определяется местом организма (популяции, вида) в круговороте. Это заявление не имеет оттенка телеологичности, ибо с позиций С + Э подхода хорошо известно, что движущая сила развития — поток энергии — имеет физическую природу (а значит, и не имеет каких-либо целей).
Для оценки количественных аспектов изменения формы во времени требуется привлечение информационного подхода, т.е. использование С+Э+И концепции. При этом субстратный подход определит преемственность и модификации структур, энергетический — одну из главных функций, а информационный — скорости перестроек и развития. Можно надеяться, что на этом пути мы сумеем хотя бы в целом вычислять и прогнозировать эволюцию формы и функции и не только на малых отрезках времени.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ВОРИШКИ, НАХЛЕБНИКИ И ПАРАЗИТЫ
ВОРИШКИ, НАХЛЕБНИКИ И ПАРАЗИТЫ Если бы мы вели себя сходным образом, мы жили бы совсем, как Алиса в Стране Чудес. Мы с восхищением содержали бы в своих домах дикобразов, аллигаторов, ракообразных и прочих тварей, настаивая, чтобы они сидели с нами за одним столом, и
Эти «примитивные» губки…
Эти «примитивные» губки… Идея «клеточного государства», в общем, довольно спорная, если речь идет о принципах строения высших животных, не покажется, вероятно, столь уж абсурдной, если мы спустимся на несколько ступенек вниз по эволюционной лестнице. Давайте задержимся
Альтруисты процветают благодаря статистическому парадоксу
Альтруисты процветают благодаря статистическому парадоксу Могут ли быть в природе ситуации, когда альтруисты ни прямо, ни косвенно не получают никакой выгоды от своего альтруизма и совсем не умеют бороться с обманщиками, но альтруизм тем не менее развивается и
Наши внутренние паразиты Эндогенные ретровирусы
Наши внутренние паразиты Эндогенные ретровирусы Идея о том, что гены носителей могли зародиться в вирусах, кажется из-за своей странности почти философской. Мы привыкли считать свой геном основой нашей личности. Мы знаем, кто является нашими биологическими родителями,
Глава 10 ПРИМИТИВНЫЕ ФОРМЫ ВРЕМЕННЫХ СВЯЗЕЙ
Глава 10 ПРИМИТИВНЫЕ ФОРМЫ ВРЕМЕННЫХ СВЯЗЕЙ Приспособительная изменчивость поведения свойственна всем живым существам. Уже простейшие могут образовывать своеобразные временные внутриклеточные связи. С появлением у кишечнополостных нервной системы (органа
Паразиты кожи
Паразиты кожи [Паразитарные заболевания кожи могут быть как растительного, так и животного происхождения. К растительным паразитам относятся только немногие плесневые грибки, к животным же паразитам, поселяющимся на коже и в ея кожице, относятся блохи, вши, власоеды,
Растительные паразиты Парша и лишай. Favus et Herpes tonsurans
Растительные паразиты Парша и лишай. Favus et Herpes tonsurans Как парша, так и лишай, вызываемые плесневыми грибками, родственными друг другу, у собак наблюдаются сравнительно редко, а поэтому играют второстепенную роль среди кожных заболеваний собак, хотя обе эти болезни легко
Животные паразиты Блохи. Aphaniptera
Животные паразиты Блохи. Aphaniptera [В Европе на коже собак паразитирует обыкновенная собачья блоха (Pulex canis), но в тропических странах (Африка и Америка), кроме этой блохи на собак, также как на птиц и на человека нападает другая, более вредная, проникающая блоха (Sarcopsylla penetrans),
Паразиты
Паразиты Обычно бембекс, прилетев с мухой к норке, без задержек опускается к порогу жилья. Но иной раз он долго парит над песком и потом спускается медленно, с жалобным жужжанием. Эта осторожность может вызвать предположение, что бембекс с высоты приглядывается, чтобы
Насекомые парализаторы и паразиты
Насекомые парализаторы и паразиты ...Мы сидим возле машины вокруг тента, постеленного на землю, завтракаем, обсуждаем маршрут путешествия. Пожалуй, нам больше не стоит задерживаться в обширной и горячей пустыне и следует ехать дальше. Таково мнение большинства. Вдруг
Паразиты и кровососы
Паразиты и кровососы Громадная армия насекомых питается кровью позвоночных животных. Многочисленные комары, москиты, слепни, мошки, мокрецы, кровососущие мухи — алчные потребители крови, досаждают животным и человеку, несут массу невзгод и мучений. В природе
Замечательные паразиты
Замечательные паразиты Мы рассказали о насекомых-хищниках. Поговорим теперь о насекомых-паразитах.…Река Чилик, молочно-белая, шумливая, бежит через Сюгатинскую долину. Она разлилась многочисленными протоками, разрезала большой зеленый тугай на множество островков и,
Глава 8. Паразиты кожи и некоторые вызываемые ими болезни
Глава 8. Паразиты кожи и некоторые вызываемые ими болезни Клещи Иксодовые, или пастбищные клещи. Временные паразиты собак. Встречаются во всех частях света, но в основном в лесной, лесостепной и степной зонах. В СССР известно более 60 видов иксодовых клещей. Многие виды
Паразиты окаянные!
Паразиты окаянные! Пухоеды похожи на вшей, но кровь не сосут, а кормятся перхотью, перьями, волосами, роговым слоем кожи, струпьями, некоторые, впрочем, и кровью, но не сосут ее, а «едят». Глотают капельки крови, вытекающие из ранок. На людях они не паразитируют; лишь на
Феминизаторы и мужские убийцы: симбионты и паразиты
Феминизаторы и мужские убийцы: симбионты и паразиты Сравнительно недавно были описаны и вовсе поразительные феномены детерминации пола у целого ряда видов беспозвоночных (насекомых, ракообразных, изопод) и механизмы контроля соотношения полов в популяции. Оказалось,