6.2. Энергетический принцип экстенсивного развития (ЭПЭР)
6.2. Энергетический принцип экстенсивного развития (ЭПЭР)
Обозначим через Нпад поток падающей энергии, т. е. поток энергии, который потенциально может быть использован некоторой биологической системой. Примерами Нпад являются поток солнечной энергии для автотрофов, количество органической пищи (в калориях), доступной для использования гетеротрофными звеньями в единицу времени. Не весь падающий поток энергии может быть захвачен и использован системой:
Нисп = Нпад — Ннеисп. (1)
Сам поток энергии, захваченной системой, складывается из двух частей: доли, связанной с запасением энергии в биомассе — В, и потерь на организацию, поддержание и активность, т. е. расход энергии системой, — R:
Нисп = µ В — R[2]. (2)
где µ — показатель прироста биомассы (или обновления в стационарном состоянии открытой системы).
Объединив (1) и (2), получим общее соотношение для потоков энергии в систему и ее трансформации в этой системе:
Нпад — Ннеисп = Нисп = µ В + R (3)
Дадим формулировку энергетического принципа: в процессах развития надорганизменных систем (эволюции, экологических сукцессиях и перестройках) использованный биологической системой поток энергии Нисп возрастает, достигая локальных максимальных значений в стационарных состояниях.
Подчеркнем еще раз, что в формулировке данного принципа речь идет о стационарных состояниях, которые достигаются в процессе развития открытых биологических систем популяционного и более высоких уровней структурной организации, а также их эволюции.
Представим схематически наиболее предпочтительный тип изменения потоков энергии во времени, использованных системой (рис. 10,а). Здесь показаны и рост Нпад, т. е. рост захваченной энергии, и уменьшение Ннеисп, т. е. снижение потерь. Штриховкой обозначено, что при этом происходит увеличение рассеяния энергии внутри системы Д с уменьшением трат на образование биомассы (это относится уже ко второму энергетическому принципу и будет обсуждаться позже). Естественно, что монотонное линейное увеличение Нпад или снижение Ннеисп не может иметь места в реальных системах. Гораздо типичнее скачкообразные переходы, связанные с качественными изменения в самой системе (рис. 10, б—г). Условия роста (или по крайней мере не убывания) потока использованной энергии Нисп остаются для каждого момента времени.
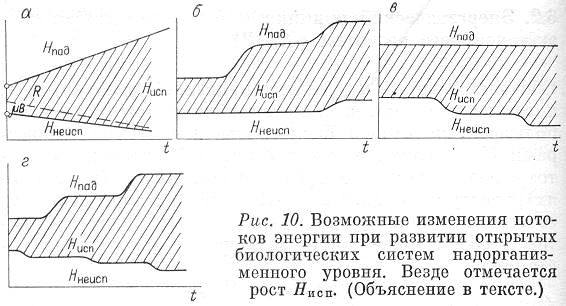
Рис. 10. Возможные изменения потоков энергии при развитии открытых биологических систем надорганизменного уровня. Везде отмечается рост Нисп. (Объяснение в тексте)
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
6.4. Энергетический принцип интенсивного развития (ЭПИР)
6.4. Энергетический принцип интенсивного развития (ЭПИР) Говоря об ЭПЭР, мы подчеркивали возрастание способности живой системы захватывать энергию, способности к экспансии, распространению в новые места. При этом качественных изменений энергетики организмов можно и не
Глава 11. Что дает энергетический подход для теоретической биологии, или попытка ответить на вопросы «почему»?
Глава 11. Что дает энергетический подход для теоретической биологии, или попытка ответить на вопросы «почему»? Многие вещи нам не понятны не потому, что наши понятия слабы, но потому, что сии вещи не входят в круг наших понятий. Козьма
Глава 2. Что такое энергетический обмен?
Глава 2. Что такое энергетический обмен? Как клетка получает и использует энергию Чтобы жить, надо работать. Эта житейская истина вполне приложима к любым живым существам. Все организмы: от одноклеточных микробов до высших животных и человека — непрерывно совершают
Принцип Митчела
Принцип Митчела В хронологии своих встреч с Митчелом я обнаружил правильную периодичность: познакомившись в 1966-м, мы затем виделись раз в два года, по четным годам. Лишь после 1974 года наступил четырехлетний перерыв, и автора хемиосмотической гипотезы я повстречал вновь
5.3. Принцип естественного равновесия
5.3. Принцип естественного равновесия Принцип равновесия играет в живой природе огромную роль. Равновесие существует между видами, и смещение его в одну сторону, скажем, уничтожение хищников, может привести к исчезновению жертв, которым не будет хватать пищи. Естественное
ПРИНЦИП ЭТОГО ОПРЕДЕЛЕНИЯ
ПРИНЦИП ЭТОГО ОПРЕДЕЛЕНИЯ К счастью, сравнительная анатомия обладала принципом, который, будучи хорошо развит, мог устранить все затруднения. Это принцип корреляции форм у организованных существ; с его помощью каждое существо могло быть в крайнем случае распознано по
Глава 10. Энергетический обмен. Биологическое окисление
Глава 10. Энергетический обмен. Биологическое окисление Живые организмы с точки зрения термодинамики – открытые системы. Между системой и окружающей средой возможен обмен энергии, который происходит в соответствии с законами термодинамики. Каждое органическое
Энергетический обмен в нервной ткани
Энергетический обмен в нервной ткани Характерными чертами энергетического обмена в ткани головного мозга являются:1. Высокая его интенсивность в сравнении с другими тканями.2. Большая скорость потребления кислорода и глюкозы из крови. Головной мозг человека, на долю
3.6.1. Принцип доминанты
3.6.1. Принцип доминанты А. А. Ухтомский создал учение о доминанте (от латинского dorninantus = господство) — временном преобладании возбуждения в нервных центрах, функционально объединяющихся для осуществления определенной деятельности. Очаг возбуждения как бы притягивает к
Принцип Оккама
Принцип Оккама Из принципа Оккама не следует, что самое простое объяснение – обязательно самое верное. Появление человека проще всего объяснить актом творения. Но правильной является эволюционная теория. Принцип Оккама означает, что к более сложным объяснениям следует
5. Теория стадийного развития и особенности развития животных
5. Теория стадийного развития и особенности развития животных В основе управления развитием организмов лежит теория стадийного развития, которую сформулировал академик Т. Д. Лысенко, исходя из работы И. В. Мичурина и многочисленных собственных исследований.Несмотря на
Принцип отдельных признаков
Принцип отдельных признаков У Г. Менделя был один удивительный предшественник, почти вскрывший основные закономерности наследования признаков за 35 лет до работ Менделя. Это О. Сажре — французский растениевод (1763–1851), автор замечательных работ по гибридизации тыквенных,