Глава 10. Энергетический обмен. Биологическое окисление
Глава 10. Энергетический обмен. Биологическое окисление
Живые организмы с точки зрения термодинамики – открытые системы. Между системой и окружающей средой возможен обмен энергии, который происходит в соответствии с законами термодинамики. Каждое органическое соединение, поступающее в организм, обладает определенным запасом энергии (Е). Часть этой энергии может быть использована для совершения полезной работы. Такую энергию называют свободной энергией (G). Направление химической реакции определяется значением DG. Если эта величина отрицательна, то реакция протекает самопроизвольно. Такие реакции называются экзергоническими. Если DG положительно, то реакция будет протекать только при поступлении свободной энергии извне – это эндергонические реакции. В биологических системах термодинамически невыгодные эндергонические реакции могут протекать лишь за счет энергии экзергонических реакций. Такие реакции называют энергетически сопряженными.
Важнейшей функцией многих биологических мембран служит превращение одной формы энергии в другую. Мембраны, обладающие такими функциями, называются энергопреобразующими. Любая мембрана, выполняющая энергетическую функцию, способна к превращению химической энергии окисляемых субстратов или АТФ в электрическую энергию, а именно в трансмембранную разность электрических потенциалов (DY) или в энергию разности концентраций веществ, содержащихся в разделенных мембраной растворах, и наоборот. Среди энергопреобразующих мембран, имеющих наибольшее значение, можно назвать внутреннюю мембрану митохондрий, внешнюю цитоплазматическую мембрану, мембраны лизосом и комплекса Гольджи, саркоплазматический ретикулум. Наружная мембрана митохондрий и ядерная мембрана не может превращать одну форму энергии в другую.
Преобразование энергии в живой клетке описывается следующей общей схемой:
Энергетические ресурсы ? ??I ? работа
где ??I – трансмембранная разность электрохимических потенциалов иона I. Следовательно, процессы утилизации энергии и совершения за счет нее работы оказываются сопряжены через образование и использование ??I. Поэтому данный ион может быть назван сопрягающим ионом. Основным сопрягающим ионом в клетке эукариот является Н+, и соответственно ??Н+ является основной конвертируемой формой запасания энергии. Вторым по значимости сопрягающим ионом является Na+ (??Na+). В то время как Сa2+, K+ и Cl- не используются для совершения какой-либо работы.
Биологическое окисление – это процесс дегидрирования субстрата с помощью промежуточных переносчиков водорода и его конечного акцептора. Если в роли конечного акцептора выступает кислород, процесс называется аэробным окислением или тканевым дыханием, если конечный акцептор представлен не кислородом – анаэробным окислением. Анаэробное окисление имеет ограниченное значение в организме человека. Основная функция биологического окисления – обеспечение клетки энергией в доступной форме.
Тканевое дыхание – процесс окисления водорода кислородом до воды ферментами цепи тканевого дыхания. Оно протекает по следующей схеме:
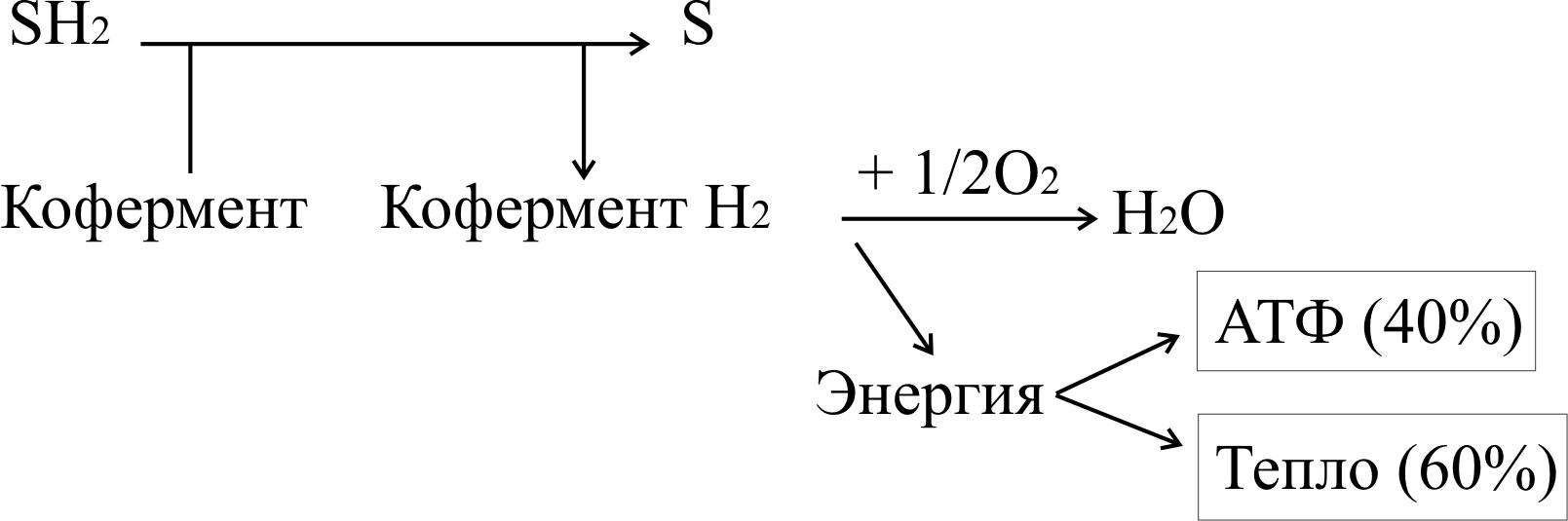
Вещество окисляется, если отдает электроны или одновременно электроны и протоны (атомы водорода), или присоединяет кислород. Способность молекулы отдавать электроны другой молекуле определяется окислительно-восстановительным потенциалом (редокс-потенциалом). Любое соединение может отдавать электроны только веществу с более высоким окислительно-восстановительным потенциалом. Окислитель и восстановитель всегда образуют сопряженную пару.
Выделяют 2 типа окисляемых субстратов:
1. Пиридинзависимые – спиртовые или альдегидные – изоцитрат, ?-кетоглутарат, пируват, малат, глутамат, ?-гидроксиацил-КоА, ?-гидроксибутират, – в их дегидрировании участвуют НАД-зависимые дегидрогеназы.
2. флавинзависимые – являются производными углеводородов – сукцинат, ацил-КоА, глицерол-3-фосфат, холин – при дегидрировании передают водород на ФАД-зависимые дегидрогеназы.
Цепь тканевого дыхания – последовательность переносчиков протонов водорода (Н+) и электронов от окисляемого субстрата на кислород, локализованных на внутренней мембране митохондрий.
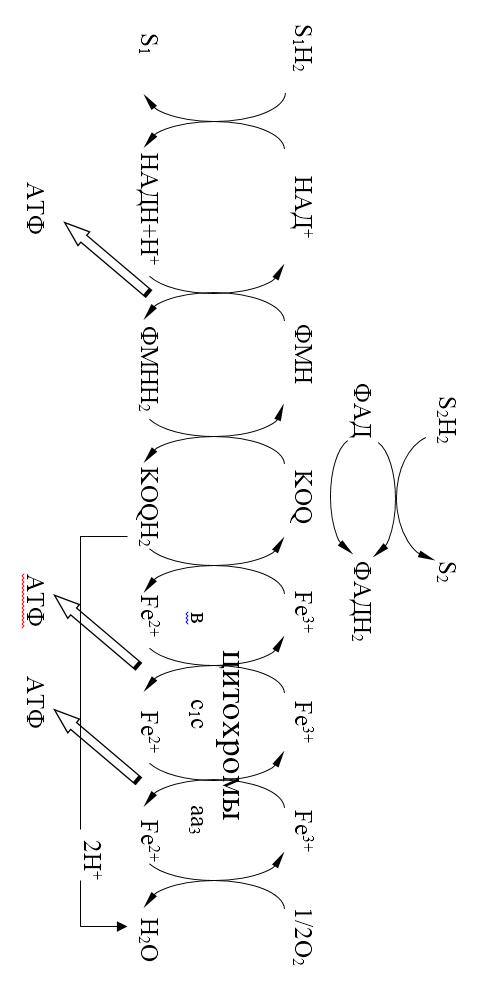
Рис. 10.1. Схема ЦТД
Компоненты ЦТД:
1. НАД-зависимые дегидрогеназы дегидрируют пиридинзависимые субстраты и акцептируют 2? и один Н+.
2. ФАД (ФМН) - зависимые дегидрогеназы акцептируют 2 атома водорода (2Н+ и 2?). ФМН – зависимая дегидрогеназа дегидрирует только НАДН, в то время как ФАД-дегидрогеназы окисляют флавинзависимые субстраты.
3. Жирорастворимый переносчик убихинон (кофермент Q, КоQ) – свободно перемещается по мембране митохондрий и акцептирует два атома водорода и превращается в КоQH2 (восстановленная форма – убихинол).
4. Система цитохромов – переносит только электроны. Цитохромы железосодержащие белки, простетическая группа которых по структуре напоминает гем. В отличие от гема атом железа в цитохроме может обратимо переходить из двух – в трехвалентное состояние (Fe3+ + ? ? Fe2+). Это и обеспечивает участие цитохрома в транспорте электронов. Цитохромы действуют в порядке возрастания их редокс-потенциала и в дыхательной цепи располагаются следующим образом: b-с1-с-а-а3. Два последних работают в ассоциации как один фермент цитохромоксидаза аа3. Цитохромоксидаза состоит из 6 субъединиц (2 - цитохрома а и 4 - цитохрома а3). В цитохроме а3 кроме железа имеются атомы меди и он передает электроны непосредственно на кислород. Атом кислорода при этом заряжается отрицательно и приобретает способность взаимодействовать с протонами с образованием метаболической воды.
Железосерные белки (FeS) – содержат негемовое железо и участвуют в окислительно-восстановительных процессах, протекающих по одноэлектронному механизму и ассоциированы с флавопротеинами и цитохромом b.
Структурная организация цепи тканевого дыхания
Компоненты дыхательной цепи во внутренней мембране михохондрий формируют комплексы:
1. I комплекс (НАДН-КоQН2-редуктаза) – принимает электороны от митохондриального НАДН и транспортирует их на КоQ. Протоны транспортируются в межмембранное пространство. Промежуточным акцептором и переносчиком протонов и электронов являются ФМН и железосерные белки. I комплекс разделяет поток электронов и протонов.
2. II комплекс – сукцинат – КоQ - редуктаза – включает ФАД- зависимые дегидрогеназы и железосерные белки. Он транспортирует электроны и протоны от флавинзависимых субстратов на убихинон, с образованием промежуточного ФАДН2.
Убихинон легко перемещается по мембране и передает электроны на III комплекс.
3. III комплекс – КоQН2 - цитохром с - редуктаза – имеет в своем составе цитохромы b и с1, а также железосерные белки. Функционирование КоQ с III комплексом приводит к разделению потока протонов и электронов: протоны из матрикса перекачиваются в межмембранное пространство митохондрий, а электроны транспортируются далее по ЦТД.
4. IV комплекс – цитохром а - цитохромоксидаза – содержит цитохромоксидазу и транспортирует электроны на кислород с промежуточного переносчика цитохрома с, который является подвижным компонентом цепи.
Существует 2 разновидности ЦТД:
1. Полная цепь – в нее вступают пиридинзависимые субстраты и предают атомы водорода на НАД-зависимые дегидрогеназы
2. Неполная (укороченная или редуцированная) ЦТД в которой атомы водорода передаются от ФАД-зависимых субстратов, в обход первого комплекса.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Глава 11. Что дает энергетический подход для теоретической биологии, или попытка ответить на вопросы «почему»?
Глава 11. Что дает энергетический подход для теоретической биологии, или попытка ответить на вопросы «почему»? Многие вещи нам не понятны не потому, что наши понятия слабы, но потому, что сии вещи не входят в круг наших понятий. Козьма
Глава 2. Что такое энергетический обмен?
Глава 2. Что такое энергетический обмен? Как клетка получает и использует энергию Чтобы жить, надо работать. Эта житейская истина вполне приложима к любым живым существам. Все организмы: от одноклеточных микробов до высших животных и человека — непрерывно совершают
Перекисное окисление липидов (ПОЛ)
Перекисное окисление липидов (ПОЛ) Реакции ПОЛ являются свободнорадикальными и постоянно протекают в организме, также как и реакции образования АФК. В норме они поддерживаются на определенном уровне и выполняют ряд функций:1. индуцируют апоптоз (запрограммированную
Глава 16. Углеводы тканей и пищи – обмен и функции
Глава 16. Углеводы тканей и пищи – обмен и функции Углеводы входят в состав живых организмов и вместе с белками, липидами и нуклеиновыми кислотами определяют специфичность их строения и функционирования. Углеводы участвуют во многих метаболических процессах, но прежде
Глава 18. Обмен гликогена
Глава 18. Обмен гликогена Гликоген – основной резервный полисахарид в животных тканях. Он представляет собой разветвленный гомополимер глюкозы, в котором остатки глюкозы соединены в линейных участках ?-1,4-гликозидными связями, а в точках ветвления – ?-1,6- гликозидными
Глава 20. Обмен триацилглицеролов и жирных кислот
Глава 20. Обмен триацилглицеролов и жирных кислот Приём пищи человеком происходит иногда со значительными интервалами, поэтому в организме выработались механизмы депонирования энергии. ТАГ (нейтральные жиры) – наиболее выгодная и основная форма депонирования энергии.
Глава 21. Обмен сложных липидов
Глава 21. Обмен сложных липидов К сложным липидам относят такие соединения, которые, помимо липидного, содержат и нелипидный компонент (белок, углевод или фосфат). Соответственно существуют протеолипиды, гликолипиды и фосфолипиды. В отличие от простых липидов,
Глава 23. Обмен аминокислот. Динамическое состояние белков организма
Глава 23. Обмен аминокислот. Динамическое состояние белков организма Значение аминокислот для организма в первую очередь заключается в том, что они используются для синтеза белков, метаболизм которых занимает особое место в процессах обмена веществ между организмом и
Глава 26. Обмен нуклеотидов
Глава 26. Обмен нуклеотидов Практически все клетки организма способны к синтезу нуклеотидов (исключение составляют некоторые клетки крови). Другим источником этих молекул могут быть нуклеиновые кислоты собственных тканей и пищи, однако эти источники имеют лишь
Глава 29. Водно-электролитный обмен
Глава 29. Водно-электролитный обмен Распределение жидкости в организмеДля выполнения специфических функций клеткам необходима устойчивая среда обитания, включая стабильное обеспечение питательными веществами и постоянное выведение продуктов обмена. Основу
Энергетический обмен в нервной ткани
Энергетический обмен в нервной ткани Характерными чертами энергетического обмена в ткани головного мозга являются:1. Высокая его интенсивность в сравнении с другими тканями.2. Большая скорость потребления кислорода и глюкозы из крови. Головной мозг человека, на долю
2.5. Биологическое окисление
2.5. Биологическое окисление Анализируя отдельные этапы клеточного метаболизма, всегда необходимо помнить, что он представляет собой единый, интегральный, взаимосвязанный механизм (Бохински Р., 1987). Процессы анаболизма и катаболизма происходят в клетке одновременно и
16. Обмен веществ и превращение энергии. Энергетический обмен
16. Обмен веществ и превращение энергии. Энергетический обмен Вспомните!Что такое метаболизм?Из каких двух взаимосвязанных процессов он состоит?Где в организме человека происходит расщепление большей части органических веществ, поступающих с пищей?Обмен веществ и