Глава 18. Обмен гликогена
Глава 18. Обмен гликогена
Гликоген – основной резервный полисахарид в животных тканях. Он представляет собой разветвленный гомополимер глюкозы, в котором остатки глюкозы соединены в линейных участках ?-1,4-гликозидными связями, а в точках ветвления – ?-1,6- гликозидными связями. Эти связи образуются примерно с каждым десятым остатком глюкозы, то есть точки ветвления в гликогене встречаются примерно через каждые десять остатков глюкозы. Так возникает древообразная структура с молекулярной массой 105 – 108 Да и выше. При полимеризации глюкозы снижается растворимость образующейся молекулы гликогена и, следовательно, её влияние на осмотическое давление в клетке. Это обстоятельство объясняет, почему в клетке депонируется гликоген, а не свободная глюкоза.
После приема пищи, богатой углеводами, запас гликогена в печени может составлять примерно 5 % от её массы. В мышцах запасается около 1 % гликогена, однако масса мышечной ткани значительно больше и поэтому общее количество гликогена в мышцах приблизительно в 2 раза больше, чем в печени. Гликоген может синтезироваться во многих клетках, например в нейронах, макрофагах, адипоцитах, но содержание его в этих тканях незначительно. В организме может содержаться до 400 г гликогена. Распад гликогена печени служит в основном для поддержания уровня глюкозы в крови в постабсорбтивном периоде. Поэтому содержание гликогена печени служит в основном для поддержания уровня глюкозы в крови в постабсорбтивном периоде. Поэтому содержание гликогена в печени изменяется в зависимости от режима питания. Гликоген мышц служит резервом глюкозы – источника энергии при мышечном сокращении. Мышечный гликоген не используется для поддержания уровня глюкозы в крови.
Синтез гликогена (гликогеногенез)
Гликоген синтезируется в период пищеварения (через 1–2 часа после приема углеводный пищи). Синтез гликогена из глюкозы, как и любой анаболический процесс, является эндергоническим, т. е. требует затрат энергии.
Синтез гликогена включает 4 этапа:
1. Фосфорилирование глюкозы до глюкозо-6-фосфата при участии гексокиназы или глюкокиназы.
2. Активация первого углеродного атома с образованием активной формы – УДФ – глюкозы.
3. Образование ?-1,4-гликозидных связей. В присутствии «затравки» гликогена (молекулы, включающей не менее 4 остатков глюкозы) фермент гликогенсинтаза присоединяет остатки глюкозы из УДФ-глюкозы к С4-атому концевого остатка глюкозы в гликогене, образуя ?-1,4-гликозидную связь.
4. Образование ?-1,6-гликозидных связей (точки ветвления молекулы). Образование их осуществляется амилозо-1,4 ? 1,6-трансглюкозидазой (ветвящий или бранчинг фермент). Когда длина линейного участка цепи включает минимально 11 остатков глюкозы, этот фермент переносит фрагмент (1 ? 4) цепи с минимальным количеством 6 остатков глюкозы на соседнюю цепь или на несколько участков глюкозы дальше, образуя ?-1,6-гликозидную связь. Таким образом, образуется точка ветвления. Ветви растут путем последовательного присоединения (1–4)-глюкозильных единиц и дальнейшего ветвления.
Гликогенсинтаза – регуляторный фермент, существующий в двух формах:
1. – дефосфорилированной, активной (форма а);
2. – фосфорилированной, неактивной (форма b).
Активная форма образуется из неактивной под действием фосфатазы гликогенсинтазы при дефосфорилировании. Превращение активной формы в неактивную происходит при участии протеинкиназы путем фосфорилирования за счет АТФ.
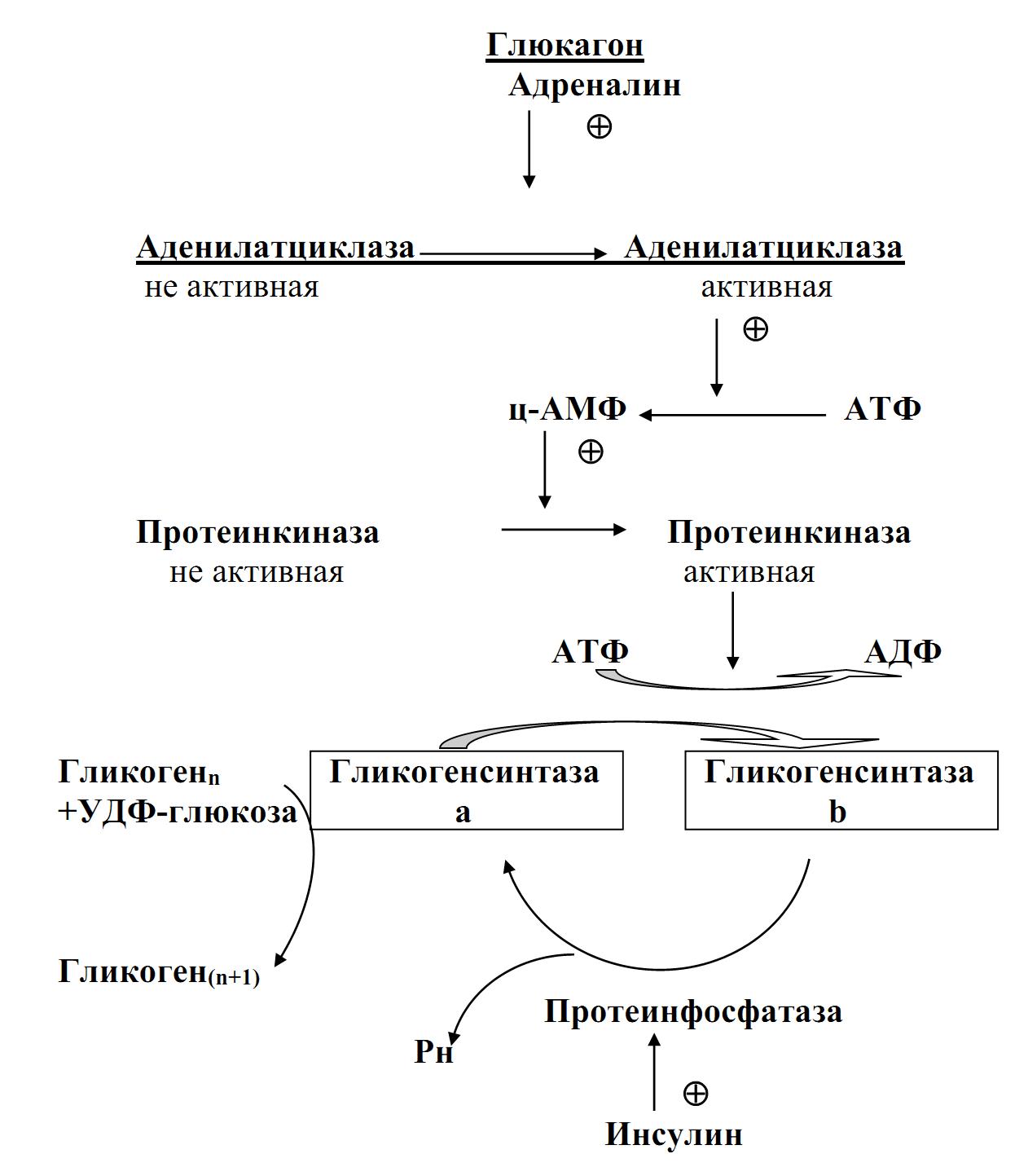
Рис. 18.-1. Регуляция активности гликогенсинтазы.
Распад гликогена может проходить двумя путями.
1. Гидролитический – при участии амилазы с образованием декстринов и даже свободной глюкозы.
2. Фосфоролитический – под действием фосфорилазы и образованием глюкозо-1-фосфата. Это основной путь распада гликогена.
Фосфорилаза – сложный регуляторный фермент, существующий в двух формах – активной и неактивной. Активная форма (фосфорилаза а) – это тетрамер, в котором каждая субъединица соединена с остатком ортофосфата через гидроксильную группу серина. Под действием фосфатазы фосфорилазы происходит дефосфорилирование, отщепление 4 молекул фосфорной кислоты, и фосфорилаза а превращается в неактивную форму – фосфорилазу b, распадаясь на две димерные молекулы. Фосфорилаза b активируется путем фосфорилирования остатков серина за счет АТФ под действием фермента киназы фосфорилазы. В свою очередь этот фермент также существует в двух формах. Активная киназа фосфорилазы – фосфорилированный фермент, превращается в неактивную форму под действием фосфатазы. Активация киназы фосфорилазы осуществляется путем фосфорилирования за счет АТФ в присутствии ионов Mg2+ протеинкиназой.
Регуляция синтеза и распада гликогена носит каскадный характер и происходит путем химической модификации ферментов.
Поскольку синтез и распад гликогена протекают по разным метаболическим путям, эти процессы могут контролироваться реципрокно. Влияние гормонов на синтез и распад гликогена осуществляется путем изменения в противоположных направлениях активности двух ключевых ферментов: гликогенсинтазы и гликогенфосфорилазы с помощью их фосфорилирования и дефосфорилирования. Инсулин стимулирует синтез гликогена и тормозит распад, адреналин и глюкагон обладают противоположным эффектом.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Глава 2. Что такое энергетический обмен?
Глава 2. Что такое энергетический обмен? Как клетка получает и использует энергию Чтобы жить, надо работать. Эта житейская истина вполне приложима к любым живым существам. Все организмы: от одноклеточных микробов до высших животных и человека — непрерывно совершают
Глава 14 Homo erectus. Развитие мозга. Зарождение речи. Интонации. Речевые центры. Глупость и ум. Смех—плач, их происхождение. Обмен информацией в группе.
Глава 14 Homo erectus. Развитие мозга. Зарождение речи. Интонации. Речевые центры. Глупость и ум. Смех—плач, их происхождение. Обмен информацией в группе. Homo erectus оказался весьма пластичным «прачеловеком»: за миллион с лишним лет своего существования он все время
Обмен веществ
Обмен веществ Наши болезни все те же, что и тысячи лет назад, но врачи подыскали им более дорогие названия. Народная мудрость — Повышенный уровень холестерина может наследоваться — Ранняя смертность и гены ответственны за утилизацию холестерина — Наследуется ли
Глава 10. Энергетический обмен. Биологическое окисление
Глава 10. Энергетический обмен. Биологическое окисление Живые организмы с точки зрения термодинамики – открытые системы. Между системой и окружающей средой возможен обмен энергии, который происходит в соответствии с законами термодинамики. Каждое органическое
Глава 16. Углеводы тканей и пищи – обмен и функции
Глава 16. Углеводы тканей и пищи – обмен и функции Углеводы входят в состав живых организмов и вместе с белками, липидами и нуклеиновыми кислотами определяют специфичность их строения и функционирования. Углеводы участвуют во многих метаболических процессах, но прежде
Синтез гликогена (гликогеногенез)
Синтез гликогена (гликогеногенез) Гликоген синтезируется в период пищеварения (через 1–2 часа после приема углеводный пищи). Синтез гликогена из глюкозы, как и любой анаболический процесс, является эндергоническим, т. е. требует затрат энергии.Синтез гликогена включает 4
Нарушения обмена гликогена
Нарушения обмена гликогена Гликогеновые болезни – группа наследственных нарушений в основе которых лежит снижение или отсутствие активности ферментов, катализирующих реакции синтеза или распада гликогена. К данным нарушениям относятся гликогенозы и
Глава 20. Обмен триацилглицеролов и жирных кислот
Глава 20. Обмен триацилглицеролов и жирных кислот Приём пищи человеком происходит иногда со значительными интервалами, поэтому в организме выработались механизмы депонирования энергии. ТАГ (нейтральные жиры) – наиболее выгодная и основная форма депонирования энергии.
Обмен кетоновых тел
Обмен кетоновых тел При голодании, длительной физической нагрузке и в случаях, когда клетки не получают достаточного количества глюкозы (желудочно-кишечные расстройства у детей, диета с низким содержанием углеводов, почечная глюкозурия, сахарный диабет), в жировой ткани
Глава 21. Обмен сложных липидов
Глава 21. Обмен сложных липидов К сложным липидам относят такие соединения, которые, помимо липидного, содержат и нелипидный компонент (белок, углевод или фосфат). Соответственно существуют протеолипиды, гликолипиды и фосфолипиды. В отличие от простых липидов,
Глава 23. Обмен аминокислот. Динамическое состояние белков организма
Глава 23. Обмен аминокислот. Динамическое состояние белков организма Значение аминокислот для организма в первую очередь заключается в том, что они используются для синтеза белков, метаболизм которых занимает особое место в процессах обмена веществ между организмом и
Глава 26. Обмен нуклеотидов
Глава 26. Обмен нуклеотидов Практически все клетки организма способны к синтезу нуклеотидов (исключение составляют некоторые клетки крови). Другим источником этих молекул могут быть нуклеиновые кислоты собственных тканей и пищи, однако эти источники имеют лишь
Глава 29. Водно-электролитный обмен
Глава 29. Водно-электролитный обмен Распределение жидкости в организмеДля выполнения специфических функций клеткам необходима устойчивая среда обитания, включая стабильное обеспечение питательными веществами и постоянное выведение продуктов обмена. Основу
Обмен железа
Обмен железа В организме взрослого человека содержится 3–4 г железа, из этого количества около 3,5 г находится в плазме крови. Гемоглобин эритроцитов содержит примерно 68 % всего железа организма, ферритин – 27 % (резервное железо печени, селезенки, костного мозга), миоглобин
ОБМЕН УГЛЕВОДОВ
ОБМЕН УГЛЕВОДОВ Следует еще раз подчеркнуть, что процессы, происходящие в организме, представляют собой единое целое, и только для удобства изложения и облегчения восприятия рассматриваются в учебниках и руководствах в отдельных главах. Это относится и к разделению на
16. Обмен веществ и превращение энергии. Энергетический обмен
16. Обмен веществ и превращение энергии. Энергетический обмен Вспомните!Что такое метаболизм?Из каких двух взаимосвязанных процессов он состоит?Где в организме человека происходит расщепление большей части органических веществ, поступающих с пищей?Обмен веществ и