Глава 7 АНАЛИТИКО-СИНТЕТИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ МОЗГА
Глава 7
АНАЛИТИКО-СИНТЕТИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ МОЗГА
Вся высшая нервная деятельность состоит из непрерывного анализа — расчленения раздражителей окружающего мира на все более простые их элементы, и синтеза — обратного слияния этих элементов в целостное восприятие действительности. Тонкая аналитико-синтетическая работа мозга обеспечивает гибкую ориентировку животного в сложной обстановке и правильное отражение окружающей действительности в сознании человека.
Генерализация и специализация условных рефлексов
Анализ и синтез всегда взаимосвязаны. Тенденция к этому проявляется в любом условном рефлексе. В результате условный рефлекс непрерывно совершенствуется. Возникая приблизительным и расплывчатым, он постепенно становится точным и определенным. Такое совершенствование условного рефлекса связано с преобладанием сначала его генерализации, а затем специализации.
Стадия генерализации. Явление генерализации особенно наглядно продемонстрировано на примере классического слюнного условного рефлекса, выработанного на кожное раздражение (касалкой).
Вдоль всего тела собаки от передней лапы через туловище до задней лапы было расставлено семь касалок, но рефлекс вырабатывали только на одну из них: уставленную на бедре. Когда рефлекс на эту касалку был выработан, попробовали остальные касалки, действие которых никогда не сочеталось с едой и ранее не вызывало слюноотделения. Однако теперь они тоже стали вызывать слюноотделение. Это не случайно: каждый новый условный рефлекс вначале имеет приблизительный, обобщенный характер.
Стадия генерализации условного рефлекса проявляется в условных ответах на все раздражители, сходные с основным сигналом.
Генерализация условного пищевого рефлекса волчонка, который впервые отведал свежей крови, поймав выпавшего из гнезда птенца, проявляется в безрассудной отваге, с которой он бросается теперь преследовать все живое, движущееся. Генерализация условного оборонительного рефлекса заставляет пуганую ворону куста бояться.
Явления генерализации условных рефлексов очень часто встречаются в повседневной жизни человека. В научном познании приблизительный, обобщенный характер первых сведений о предмете всегда предшествует достижению точных знаний.
Физиологический механизм и биологическое значение генерализации. Для того чтобы понять физиологический механизм генерализации условного рефлекса, необходимо вспомнить главное условие образования временных связей. Это условие заключается в совпадении во времени возбужденного состояния пунктов сигнала и подкрепления. Следовательно, если близкие к основному другие сигналы тоже оказались связанными с подкрепляющей реакцией, очевидно, соответствующие им пункты были возбуждены, когда протекало возбуждение основного сигнального пункта. Это происходит благодаря свойству иррадиации возбудительного процесса.
О ходе нервных процессов, происходящих в мозге при генерализации условного рефлекса, ясное представление дает рис. 48. Основным очагом возбуждения при действии сигнала здесь был пункт проекции бедренной касалки в кожном анализаторе. Однако с пищевым подкреплением оказались связанными также пункты проекции касалок голени, туловища и даже передней лапы. Как это могло произойти, схематически показано на рис. 48.
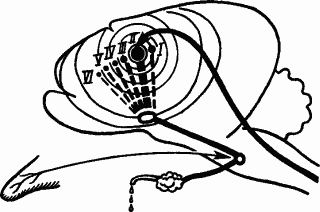
Рис. 48. Иррадиация возбуждения при образовании побочных временных связей и генерализации условного рефлекса:
I, II, III, IV, V, VI — пункты проекции в кожном анализаторе участков кожи под касалками, расположенными последовательно по задней конечности и туловищу; условный рефлекс вырабатывается на касалку II, расположенную на бедре; концентрическими кругами от пункта II изображена иррадиация возбуждения
Возбуждение, возникшее в пункте II, иррадиировало по всему кожному анализатору. Поэтому к моменту подкрепления в возбужденном состоянии кроме пункта бедра (II) оказались и пункт конца задней лапы (I), и пункт таза (III), и пункт туловища (IV) и др. Эти пункты «самопроизвольно» без специальной выработки образовали побочные временные связи с пищевым подкрепляющим рефлексом.
Таким образом, физиологический механизм стадии генерализации условного рефлекса заключается в образовании побочных временных связей подкрепляющей реакции с пунктами сигналов, близких к основному. Эти пункты получили возбуждение в результате его иррадиации из пункта основного сигнала. Следовательно, в первоначальном свойстве сигналов, близких к основному, и генерализации условных рефлексов проявляются уже известные свойства движения нервных процессов в высших отделах мозга.
Каково биологическое значение генерализованного характера новых условных рефлексов? Выгоду генерализованности новых условных рефлексов можно оценить на примере упомянутого выше волчонка, который поймал выпавшего из гнезда птенца. Если представить, что условный пищевой рефлекс при этом у него образовался бы не генерализованным, а сразу очень точным, то волчонок впредь охотился бы только на птенцов и оставался равнодушным к другой пище. Именно генерализация этого новообразованного охотничьего условного рефлекса заставляет его преследовать все живое, движущееся. Не меньшее значение имеет первоначальная генерализация в образовании оборонительных условных рефлексов. Именно благодаря страху перед каждым незнакомым предметом «пуганая ворона» может избежать опасности быть подстреленной.
С явлением генерализации связаны некоторые процессы обобщения в мыслительной деятельности человека.
Стадия специализации. Можно вновь рассмотреть опыт с выработкой условного пищевого рефлекса слюноотделения. Если продолжать укрепление основного рефлекса на касалку с бедра, а остальные касалки применять, не сопровождая кормлением, то через некоторое время основной рефлекс на касалку с бедра становится прочным, а рефлексы с других касалок начинают быстро ослабевать. Затем они совершенно исчезают, и остается основной, и теперь единственный, точный и определенный условный рефлекс на касалку с бедра. Рефлекс стал специализированным. Стадия специализации условного рефлекса проявляется в том, что условные ответы может вызывать только основной сигнал. Все сходные с ним теряют свое сигнальное значение, приобретенное в стадию генерализации.
Так, волчонок после легкой победы над птенцом будет преследовать каждую встречную птицу. Но вскоре опыт безуспешных попыток схватить улетающих взрослых птиц заставит его бросить это напрасное занятие и сосредоточить внимание на поисках доступных гнезд — его охотничий условный рефлекс специализировался. «Пуганая ворона» со временем перестанет бояться не только безобидного куста, но даже человека.
В жизни человека первые впечатления, в силу своеобразной генерализации мысли, иногда бывают преувеличенными. Более глубокое знакомство с вопросом позволяет укрепить то, что соответствует действительности, и отбросить неподкрепляемое фактами.
Физиологический механизм и биологическое значение специализации. При специализации условных рефлексов основной сигнал подкрепляется, смежные не подкрепляются, т.е. возникают условия для развития дифференцировочного торможения. В результате основная временная связь усиливается, а побочные ослабевают. Следовательно, физиологический механизм стадии специализации условного рефлекса заключается в угашении побочных временных связей на смежные раздражители благодаря развитию дифференцировочного торможения. Условное возбуждение на основной сигнал концентрируется точно в его анализаторном пункте. Схематическое изображение этого механизма в случае описанного выше опыта с условным слюноотделительным рефлексом с кожного анализатора представлено на рис. 49.
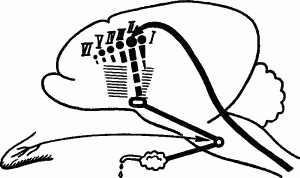
Рис. 49. Концентрация возбуждения при угашении побочных временных связей и специализации условного рефлекса:
обозначения те же, что в рис. 48, временная связь пищевой реакции с пунктом касалки на бедре (II) укреплена, побочные связи со всеми другими пунктами угашены дифференцировочным торможением, штриховкой изображена зона торможения
Что касается биологического значения специализации условного рефлекса, то оно не требует особых пояснений. Если продолжить историю приведенных выше примеров, то в результате специализации условных рефлексов волчонок научится правильно выбирать объекты своей охоты, «пуганая ворона» сохранит жизнь, не обращаясь в бегство даже от того, что может быть источником корма. Специализация восприятия позволяет человеку уточнять свои знания.
Условные рефлексы на комплексные раздражители
В реальной жизненной обстановке возникающие сигналы никогда не бывают столь простыми и единичными, как применяемые в лаборатории. Естественные сигналы всегда сложные, составные. Вид присевшего под кустом зайца, его запах, хруст обрываемых молодых побегов — все это составляет для подкрадывающейся лисы единый сигнал для завершающего охоту прыжка. Человек узнает знакомого по чертам лица, звукам голоса, походке и другим признакам, из которых и складывается общий облик, сохраняемый нашей памятью.
Не одиночные, а комплексные раздражители являются в естественной обстановке основными условными сигналами. Они образуют условные рефлексы, несравненно более сложные и изменчивые, чем рефлексы, возникающие на одиночные сигналы. В комплексных условных рефлексах идет непрерывный синтез новых комплексов и анализ их отношений к отдельным членам. Здесь течение анализа и синтеза приближается к их действительной природной сложности. Поэтому, изучая свойства условных рефлексов на комплексные раздражители, можно узнать некоторые общие правила аналитическо-синтетической работы мозга.
Условные рефлексы на одновременные комплексы раздражений. Если у животного вырабатывать условный рефлекс на несколько одномоментно действующих раздражителей, то они объединяются в одновременный комплекс, воспринимаемый как целостный образ. При этом каждый из этих раздражителей, примененный отдельно, может вызывать у животных рефлекс, сила и вероятность проявления которого будут находиться в зависимости от силы и биологической значимости раздражителя.
Условные рефлексы на суммарные раздражители. Если применять несколько условных сигналов вместе, то они объединятся в комплекс. Такой комплекс, образованный из отдельных уже готовых условных раздражителей, называется условным суммарным раздражителем. Условный рефлекс на суммарный раздражитель значительно превышает величину каждого из составивших его рефлексов в отдельности. Последние, по мере укрепления комплекса, еще более ослабляются. При этом относительно сильные раздражители могут инактивировать более слабые. Сам комплекс выступает в роли наиболее сильного раздражителя. В опыте это проявляется следующим образом. У собаки были выработаны три пищевых условных рефлекса: на тон, вертящийся предмет и вспыхивание лампочки; испытано суммарное действие этих сигналов и определена устойчивость к условному тормозу — механическому раздражению кожи. Наиболее значительное слюноотделение проявлялось в ответ на комплекс. Преобладающая сила суммарного раздражителя наглядно обнаружилась также в способности противостоять условному тормозу, который полностью подавлял рефлексы на отдельные его компоненты.
Условные рефлексы на последовательные комплексы раздражителей. Если несколько индифферентных раздражителей, следующих друг за другом с «перекрытием» в определенном и всегда постоянном порядке, сопровождать подкрепляющей реакцией, то образуется условный рефлекс на последовательный комплекс, ряд последовательно действующих раздражителей сольется в синтетический сложный сигнал.
В последовательном комплексе наиболее сильным оказывается последний подкрепляемый сигнал, а сигнальное значение остальных членов комплекса оказывается тем большим, чем ближе они стоят к подкреплению.
Условные рефлексы на цепи раздражителей. Если индифферентные раздражители, из которых образуется комплексный сигнал, действуют один за другим без всякого «переслаивания», то возникает условный рефлекс на цепь раздражителей. В этом случае ни один из членов цепи не действует одновременно с другими, а подкрепляется только последний из них. Поэтому сигнальное, пусковое значение приобретает только последний из них, а остальные создают «условно-рефлекторную настройку» (И.П. Павлов), обеспечивая «предпусковую интеграцию» (П.К. Анохин), и усиливают действие пускового раздражителя.
Образование условных рефлексов на цепи раздражителей лежит в основе выработки различных так называемых двигательных навыков путем подкрепления случайных или вынужденных движений. Например, попавший в капкан волк может случайно надавить пружину замка и освободиться. Теперь это движение приобретает значение сигнала защитной реакции. Или, например, хозяин говорит собаке «дай лапу» и поднимает ее лапу, «награждая» животное лакомым куском. Вскоре собака, услышав эти слова, сама «подает лапу».
На рис. 50 приведена схема образования такого условного рефлекса на цепь раздражителей из звуков слова, проприоцептивных сигналов поднятия лапы и, наконец, натуральных сигналов пищи. Вначале прокладываются временные связи между тремя очагами возбуждения: слухового, двигательного и пищевого. Затем закрепляется последовательность действия членов цепи. Наконец, уточняется положение основных ее членов: звукового сигнала («дай лапу»), проприоцептивных (движение конечности) и натурального пищевого (подкармливание).

Рис. 50. Рефлекторные пути двучленного двигательного условного рефлекса (по Л.Г. Воронину):
1 — слуховые рецепторы, 2 — пути звуковых сигналов, 3 — корковая область слухового анализатора, 4 — корковый отдел пищевого центра, 5 — двигательные нервные элементы коры, 6 — подкорковые двигательные аппараты, 7 — мускулатура, 8 — центробежные пути, 9 — центростремительные пути, 10 — корковая область проприоцептивного анализатора, 11 — временные связи между корковыми очагами возбуждения центра.
Так как жизнь все время изменяет сигнальное значение членов комплексов, рефлексы на комплексные раздражители непрерывно перестраиваются, внося все новые усложнения в текущую аналитико-синтетическую работу мозга.
Целостность в условно-рефлекторной деятельности
Происходящий наряду с анализом непрерывный синтез мозговой деятельности постоянно сливает отдельные раздражители в комплексы нарастающей сложности. Эти комплексы вступают между собой в еще более сложные отношения: например, следующие друг за другом комплексы одновременных раздражителей будут синтезироваться в последовательные цепи комплексов. В конечном итоге, все раздражители внешнего окружения и внутреннего состояния организма оказываются так или иначе связанными между собой. Целостность условно-рефлекторной деятельности проявляется в системности, стереотипии, «настройках», «переключениях» реакций по сигналам обстановки. Благодаря этому поведение животных определяется не одиночными сигналами, а всей целостной картиной окружающего; благодаря этому сознание человека дает целостное и единое зеркальное отражение объективно существующего мира. В результате этого высшая нервная деятельность, охватывая многие стороны настоящего, связывает его с опытом прошлого и может осуществлять тонкое приспособление к событиям будущего.
Системность в работе мозга. Если у собаки выработать несколько условных рефлексов и всегда повторять их в строго определенном порядке, то эти рефлексы свяжутся в стройную взаимозависимую систему. Такая системность рефлексов четко выявляется в следующем опыте.
У собаки вырабатывали слюноотделительные рефлексы на свет, касалку, звонок, шипение и метроном с частотой 60 ударов/мин, от которого был отдифференцирован метроном с частотой 120 ударов/мин. Рефлексы укрепляли и испытывали через 5-минутные интервалы в постоянном порядке. Результаты представлены на рис. 51. Затем вместо поочередного вызывания этих рефлексов в прежней последовательности экспериментатор стал подавать через 5-минутные интервалы один и тот же раздражитель. Этот раздражитель действовал не только на «своем месте», но и на местах остальных сигналов, вместо них. Оказалось, что, попадая на «чужое место», каждый условный раздражитель приобретал черты действия сигнала, обычно находящегося на этом месте (рис. 51, б, в, г). Например, попадая на место дифференцировочного сигнала, все раздражители резко ослабляют свое действие. Это «тормозное место». Попадая на место сильного условного раздражителя (звонка), даже слабый раздражитель (свет) стал вызывать совершенно не свойственное ему большое слюноотделение. Мозг запечатлел порядок следования рефлексов на разные сигналы, связал их все в определенную систему.
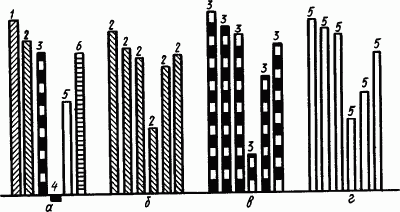
Рис. 51. Системность в работе больших полушарий головного мозга (по Э.А. Асратяну):
а — применение системы разных условных раздражителей, б — повторение одного и того же метронома (60 ударов/мин) вместо разных условных раздражителей, в — повторение шипения, г — повторение вспышек света вместо разных условных раздражителей; 1 — звонок, 2 — метроном с частотой 60 ударов/мин, 3 — шипение, 4 — метроном с частотой 120 ударов/мин, 5 — свет, 6 — касалка
Такая системность работы мозга является высшим синтезом всей совокупности действующих раздражителей окружающей среды, где каждый из них занимает определенное место во времени и пространстве.
Системность условно-рефлекторной деятельности объясняет целостность восприятия человека. Проблема целостности долгое время была предметом дискуссий и даже вызвала своеобразный кризис в психологии. Результатом этого кризиса был отказ от крайнего аналитического так называемого ассоциативного направления и возникновение другой крайности в виде так называемой гештальтной теории. Исходя из того, что целостное восприятие нельзя понять как простую сумму действующих раздражителей, но не используя учение И.П. Павлова об аналитико-синтетической деятельности мозга, психологи-гештальтисты рассматривали восприятие как изначально данные неразложимые целостные образы. Учение о системности в работе мозга раскрывает физиологический механизм, создающий целостность восприятия.
Динамический стереотип раздражителей. Системность в работе мозга проявляется тем ярче и сильнее, чем лучше выражена стереотипия раздражителей. Стереотипом раздражителей называют их набор, расположенный в строго определенном порядке в пространстве и времени и неизменно повторяющийся в той же последовательности.
Такой набор раздражителей может быть стандартным по какому-нибудь одному признаку и представлять небольшие вариации в каких-то других отношениях. Основой стереотипии становится наиболее стандартный признак. Поэтому различают виды стереотипов по качеству раздражителей, силе раздражителей, по расположению раздражителей в пространстве и по расположению их во времени. Например, лиса, как многие другие хищники, обходит свой охотничий район в определенной последовательности (стереотип в пространстве), а бродячая собака прибегает к столовой в часы обеда (стереотип во времени). Человек привыкает вставать в определенное время, идти на работу по одному и тому же пути, видеть одни и те же лица товарищей, выполнять знакомую работу.
Стереотип раздражителей в природной жизни животных отражает экологические особенности среды обитания. Стереотипы раздражителей в жизни человека определяются его привычным местом в общественной среде. При изменении условий стереотип раздражителей перестраивается, почему и называется динамическим.
Свойства и значение стереотипии рефлексов. В результате анализа и синтеза стереотипов раздражителей складываются постоянные стереотипы вызываемых ими рефлексов. Целостность восприятия дополняется целостностью поведения. Стереотипы обладают следующими важными характеристиками.
1. Стереотипные действия легче выполнять. Известно, как облегчается ходьба, когда она образует ритмический стереотип маршировки. К любой работе нужно приноровиться, и тогда она уже не требует таких усилий, как вначале.
2. Сложившийся стереотип направляет образование новых навыков по своему руслу. Например, при освоении новых объектов охоты хищник использует наиболее надежные из уже знакомых ему охотничьих приемов. Порой заключения о людях и вещах также нередко несут на себе отпечаток сложившегося стереотипа предвзятых мнений.
3. Стереотип позволяет адекватно реагировать, несмотря на некоторые изменения в обстановке. Так, однажды выработанный двигательный стереотип управления автомашиной позволяет водить машины даже с несколько различным управлением.
Из рассмотрения свойств стереотипии рефлексов можно понять ее исключительное значение в поведении животных и жизни людей. Охота хищников, норовая деятельность грызунов, пастьба травоядных, гнездостроение птиц — всюду проявляются стереотипы рефлексов. Из стереотипов комплексных рефлексов животных в природе складывается то, что называют их образом жизни. Весь уклад жизни человека ведет к формированию бесчисленного множества бытовых и трудовых стереотипов. Они проявляются возникновением аппетита в привычное время еды и в подъеме настроения после утренней зарядки, во всевозможных бытовых привычках и в поразительной точности двигательного стереотипа последовательных трудовых операций на работе. Для пунктуального человека весь распорядок дня может стать жестким стереотипом. Важной задачей педагогики является соблюдение условий, облегчающих учащемуся выработку нужных стереотипов. Это относится и к организации учебы (режим школьного дня, расписание уроков), и к содержанию занятий (объяснение дальнейшего, пользуясь усвоенным).
С возрастом стереотипы крепнут и их становится все труднее изменять. Отсюда известный консерватизм пожилых людей, у которых к тому же снижается подвижность основных нервных процессов. Переделка сложившихся стереотипов всегда представляет значительную трудность для нервной системы. Крутая ломка привычного уклада жизни может вызывать даже нервные расстройства. Люди, проработавшие всю свою жизнь и оставившие работу по старости, нередко тяжело переживают вынужденное безделье не только из-за сознания отрыва от общественно-полезной деятельности, но и из-за ломки сложившихся стереотипов поведения.
Условно-рефлекторная настройка. Очень сложные стереотипы поведения складываются с участием комплексных раздражителей обстановки. Так, например, образование последовательных комплексов из обстановочных и основных условных раздражителей по типу цепи с далеко расставленными звеньями является физиологическим механизмом так называемой условно-рефлекторной настройки. Как показывает само название — настройка, речь идет не о совершении конкретной деятельности, а о вызванном по механизму временной связи состоянии готовности к этой деятельности.
Возникновение условно-рефлекторной настройки ясно проявляется, например, если сравнивать поведение двух собак, у одной из которых вырабатывают пищевые условные рефлексы, у другой — оборонительные. Когда собаку, служащую для опытов с пищевыми рефлексами, приводят в лабораторию, она бросается навстречу экспериментатору, лижет ему руки, виляет хвостом и сама вскакивает в станок. Когда же приводят собаку, служащую для работы с оборонительными рефлексами, особенно на болевом подкреплении, то она упирается, поджимает хвост и распластывается на полу, ее приходится брать на руки, чтобы поставить в станок, откуда она пытается при первой возможности убежать.
Условно-рефлекторная настройка поведения имеет громадное приспособительное значение в жизни животных. Услышав далекий рев тигра, буйвол настораживается. Он чаще оглядывается на окружающие его заросли, внимательнее принюхивается к доносящимся запахам. Такая «оборонительная настройка» может спасти ему жизнь.
Условно-рефлекторная настройка действует и в жизни человека. Всем понятно, какое значение имеют четкость и быстрота действий, например, для солдата или спортсмена. Недаром человеческий опыт привел к широкому использованию в военном деле и спорте так называемых предварительных команд. Для бегуна на старте предварительная команда «приготовиться» является условно-рефлекторной двигательной настройкой. При этом сигнале повышается возбудимость соответствующих локомоторных механизмов, мобилизуются кровообращение, дыхание. Такая «стартовая реакция» обеспечивает готовность к немедленному напряжению всех сил в момент броска по команде. От правильности ее отработки может зависеть исход состязания.
Условно-рефлекторная настройка проявляется и в повседневных восприятиях. Художественную иллюстрацию этого мы находим, например, в рассказе К. Гельвеция о даме и священнике, смотревших в телескоп на Луну. Дама воскликнула: «Я вижу там тени двух влюбленных», священник возразил: «Вы ошиблись, сударыня, это две башни на главной колокольне».
Когда раздражители обстановки начинают играть более значительную роль в сигнальном комплексе, они могут вызвать не только настройку на определенный вид деятельности, но и самую деятельность. Характер этой деятельности при одних и тех же основных сигналах может быть иным в разной обстановке. Такое образование комплексов разного сигнального значения из одних и тех же основных сигналов с добавлением разных обстановочных раздражителей проявляется в так называемом условно-рефлекторном переключении.
Явление переключения обнаружили при выработке условных рефлексов на один и тот же сигнал, но с различным подкреплением в разной обстановке. Оно четко проявляется в следующем опыте.
Собака находилась в помещении, где звук бульканья сопровождали подкармливанием. Когда же ее приводили в другое помещение, то здесь звук бульканья сопровождали болевым раздражением лапы. Вскоре звук бульканья стал раздражителем условного рефлекса. В обстановке помещения, где бульканье сопровождалось дачей корма, оно вызывало слюноотделение. В обстановке другого помещения, где бульканье сопровождалось болевым раздражением конечности, то же бульканье вызывало отдергивание лапы. Обстановочные раздражители переключали условное возбуждение в первом случае в русло пищевой, во втором — оборонительной реакции.
Нейрофизиологический механизм переключения считают своеобразной формой тонического условного рефлекса, создающего предрасположение к определенной форме деятельности (Э.А. Асратян, 1970). Такой взгляд получил подтверждение в опытах регистрации активности нейронов гиппокампа кролика при переключении рефлекса на звук с пищевого на оборонительный, в изучении динамики ЭКоГ и ВП коры мозга собак в процессе переключения, показавших развитие при этом тонического торможения и при наблюдении за свободным поведением животных в условиях переключения действия в инструментальном пищевом или питьевом условных рефлексах.
Недавно выдвинута гипотеза информационного переключения, согласно которой в зависимости от характеристик межимпульсных интервалов, задаваемых системой оценки подкрепления, интервалселективные цепи нейронов могут менять каналы эфферентного выхода одном и том же афферентном сигнале (Г.А. Вартанян и др., 1986).
Переключения реакций по сигналам обстановки часто наблюдаются в повседневной жизни. Достаточно вспомнить поведение кошки, которая убегает от собаки во дворе и смело нападает на ту же собаку, подошедшую в комнате к корзине с котятами. Каждому человеку, конечно, приходилось испытывать переключающее действие обстановки, которая может так сильно менять направление мыслей.
Сложные формы высшей нервной деятельности животных
Приспособительное поведение животных, особенно в сложных ситуациях, нередко принимает такие формы, которые трудно «уложить» в рамки рассмотренных выше актов элементарной условно-рефлекторной деятельности. Наблюдая, как шимпанзе ставит ящик на ящик и, взобравшись на них, достает приманку, подвешенную к потолку клетки, И.П. Павлов говорил: «Когда обезьяна строит свою вышку, чтобы достать плод, то это „условным рефлексом“ назвать нельзя. Это есть случай образования знания, уловления нормальной связи вещей». Действующая при этом сложная система условных связей на основе пищевой мотивации обеспечивает целесообразную последовательность действий животного, посредством которых оно достигает удовлетворения своей потребности. Исследование нейрофизиологических механизмов таких сложных форм поведения животных составляет очередную задачу физиологии высшей нервной деятельности. Значение таких исследований определяется и тем, что раскрытие этих механизмов образования своеобразных сложных систем условных связей может пролить свет на биологические предпосылки формирования некоторых свойств аналитико-синтетической деятельности мозга человека.
Одной из таких сложных форм высшей нервной деятельности является так называемое образное или психонервное поведение (И.С. Бериташвили, 1963). Представление о своеобразии такого поведения основывается на фактах, полученных при изучении пищевых реакций животных в условиях свободного поведения. Так, достаточно однажды покормить собаку в определенном месте и она будет туда прибегать. Предполагают, что при первом же поедании корма возникают множественные связи всех признаков местонахождения пищи и ее самой в виде синтетического их образа. Этот образ хранится в памяти, и когда по внешним (обстановка) или внутренним (голод) побуждениям он репродуцируется, то осуществляет соответствующее поведение. В случае возникновения образа угрожающей ситуации репродукция этого образа по какому-либо его компоненту направляет поведение на избегание опасности.
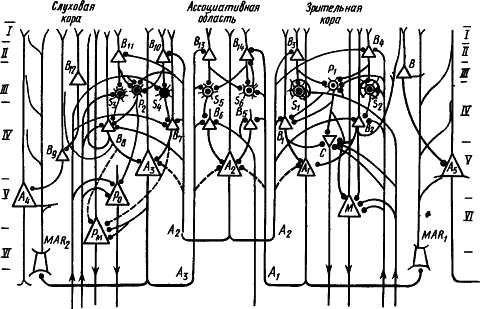
Рис. 52. Нейронный механизм объединения слухового и зрительного восприятия, создающего целостный образ звучания объекта (по И.С. Беритову):
S1 — S6 — сенсорные нейроны, P1 — P2 — передаточные звездчатые нейроны, B1 — В14 — вставочные пирамидные нейроны, A1 — A5 — ассоциативные пирамидные нейроны, РM — пирамидные проекционные нейроны, M — пирамидные нейроны Мейнерта, C — звездчатые нейроны Кахаля, MAR1 — MAR2 — вставочные нейроны Мартинотти, I—VI —слои коры; стрелками вверх и вниз обозначены афферентный вход и эфферентный выход соответственно
Считают, что в отличие от простых условных рефлексов, требующих повторения сочетаний сигнала и подкрепления, условные связи формирования образов возникают сразу. В качестве доказательства различия образного и простого условно-рефлекторного поведения приводят результаты опытов с выработкой пищевых условных рефлексов при усложненном подходе к кормушке. Когда звучал сигнальный звонок, находящийся от кормушки в 4–5 м, собаку сначала подводили к нему, потом вели в другую сторону и только затем приводили к кормушке для подкрепления едой. Оказалось, что после первого поедания корма собака стала бежать прямо к кормушке, и несмотря на сотни сочетаний условный рефлекс следования по усложненному пути был неустойчив и прерывался прямой побежкой к кормушке. Образ местонахождения пищи оказывался ведущим в поведении.
Поиски нервного субстрата формирования образов привели к заключению о том, что оно осуществляется с участием подкорковых структур совместной деятельностью проекционных и ассоциативных областей коры мозга, в которой особую роль играют звездчатые нейроны (И.С. Беритов, 1969). Их морфофизиологические особенности позволяют предположить, что звездчатые нейроны с околоклеточной сетью аксонов, образующие своеобразную матрицу в IV слое проекционных полей коры, являются сенсорными, порождающими ощущения; а звездчатые нейроны с аксонами, идущими к вставочным и ассоциативным пирамидным клеткам, связывают через них отдельные компоненты восприятия разных модальностей в целостный образ внешнего мира. На рис. 52 показан возможный нейронный механизм объединения зрительных и слуховых компонентов восприятия в целостный образ.
Другая сложная форма высшей нервной деятельности, демонстрирующая «улавливание нормальной связи вещей», о котором говорил И.П. Павлов, была исследована на примере экстраполяционного, или рассудочного, поведения (Л.В. Крушинский, 1977). Как показывает само название, речь идет о способности животного, наблюдавшего течение существенного для него события, «уловить» закономерности его протекания, а когда наблюдение прерывается, то экстраполировать, т.е. «мысленно» продолжить, ход событий и соответственно строить свое поведение без специального обучения. Примером такого поведения в природных условиях может служить случай, описанный известным натуралистом Э. Сетон-Томпсоном, когда ворона, пролетая над ручьем, уходящим в трубу, уронила в него кусок хлеба, который она несла в клюве. Заглянув внутрь, она быстро облетела трубу, дождалась выплывания куска, выловила его и полетела дальше.
Для экспериментального исследования способности к экстраполяции была разработана методика движущейся кормушки, скрывающейся на некоторое время в коридоре (Л.В. Крушинский, 1960).
На рис. 53 показана схема такой установки и результаты опытов на ней с разными птицами. Кормушка исчезала из поля зрения, проходя со скоростью 8–10 см/с через два полутораметровых коридора, между которыми оставался промежуток в 3–5 см. Как видно по поведению подопытных птиц, только сорока могла довольно точно экстраполировать движение кормушки и «предвидеть» ее появление из коридора.
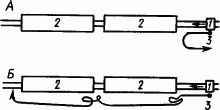
Рис. 53. Поведение голубя (А) и сороки (Б) в опыте с экстраполяцией направления движения (по Л.В. Крушинскому):
1 — кормушка, двигающаяся по рельсам в направлении, указанном стрелками, 2 — коридоры, в которых движение кормушки перестает быть видимым, 3 — путь птицы
Исследования, проведенные с помощью соответственно модифицированных методик (прохождение кормушки под горизонтальной плоскостью, помещение кормушки в движущиеся объемные фигуры и др.), позволили дать сравнительную оценку способности к экстраполяции у исследованных животных разных видов. Так, среди млекопитающих на 1-м месте оказались обезьяны, дельфины и бурые медведи, 2-е место заняли лисицы, волки, собаки, 3-е — песцы и лисицы звероферм, 4-е — кошки, 5-е — мышевидные грызуны, зайцы, 6-е — пасюки, кролики. Среди птиц на 1-м месте — вороновые, на 2-м — хищные птицы, утки, куры, на 3-м — голуби. Способность к экстраполяции у черепах оказалась на уровне между 1-м и 2-м местами у птиц. Лягушки и жабы были совершенно неспособны к обучению, рыб можно было обучить экстраполяции.
Поиски нейрофизиологических механизмов, лежащих в основе экстраполяционного поведения, привели к заключению, что у млекопитающих особую роль в его осуществлении играют лобные отделы мозга, удаление которых делает экстраполяцию невозможной. У птиц, по-видимому, эту роль выполняет особый вырост стриарных тел — вульст, развитие которого коррелирует с показателями экстраполяционной деятельности. У рептилий такая деятельность нарушается при повреждении передних отделов примитивной дорсальной коры мозга. В отношении нейронной организации экстраполяционных процессов предполагают, что они осуществляются при участии нейронов, детектирующих характер изменения действующих раздражителей, свойства которых формировались генетически в онтогенезе.
Нейрофизиологические механизмы аналитико-синтетической деятельности
Анализ воздействия окружающей среды начинается с общеизвестной избирательной чувствительности рецепторов к раздражителям различной модальности. Такой качественный анализ, позволяющий воспринимать раздельно видимое, слышимое, пахнущее и т.п., дополняет количественный анализ, когда интенсивность воздействия кодируется амплитудой рецепторного потенциала, а затем частотой импульсов в афферентном нерве. Однако уже на этом первичном нервном уровне начинаются и процессы синтеза. Одним из них является интеграция сигналов от соседних рецепторов. Это впервые было показано на сложном глазе мечехвоста (Limulus) (Г. Хартлайн, Ф. Ратлифф, 1957). Разряд импульсов от одного омматидия тормозится при освещении другого, находящегося поблизости. Благодаря такому латеральному торможению освещение обоих рецепторов дает меньший эффект, чем каждого в отдельности. Оно выделяет контуры изображения и делает его более контрастным.
В сетчатке позвоночных обнаружены более сложные формы интеграции нейронных реакций, связанные со специфическим пространственным распределением их рецептивных полей. Типичными для такого распределения являются концентрические рецептивные поля (С. Каффлер, 1953). Для них характерны противоположные реакции центра и окружения. При этом выделяются так называемые on-нейроны, которые возбуждаются точечным засветом центра своего рецептивного поля, и off-нейроны, возбуждающиеся при выключении этого засвета; нейроны, которые возбуждаются и в том и в другом случае, обозначаются как on—off-нейроны. Засвет периферии рецептивного поля вызывает противоположные реакции, т.е. on-нейроны затормаживаются, a off-нейроны возбуждаются. Тормозное взаимодействие центра и периферии рецептивного поля осуществляется горизонтальными и амакриновыми клетками. В сигналах от нейронов дискретных концентрических рецептивных полей кодируются локальные перепады освещенности частей воспринимаемого изображения.
На более высоких уровнях обработки сенсорной информации происходит дальнейший синтез афферентных сигналов о признаках объекта. В связи с разной функцией нервных клеток они обнаруживают рецептивные поля разной конфигурации. На рис. 54, А показаны ответы нейрона зрительной коры кошки на точечный засвет в центре рецептивного поля и в зоне его периферии, а также на общее освещение. По наличию реакций в первом и во втором случаях этот нейрон относится к on — off-типу. Отсутствие реакций на общий засвет указывает на уравновешенность возбуждения в on- и торможения в off-нейронах. Примечательно, что рецептивное поле этого нейрона имело характерную вытянутую в виде полосы форму (рис. 54, Б). Значение такой формы рецептивного поля раскрывается в опытах с изменением наклона световой полоски, что приводит к уменьшению реакции при перекрытии полоской периферической части рецептивного поля.
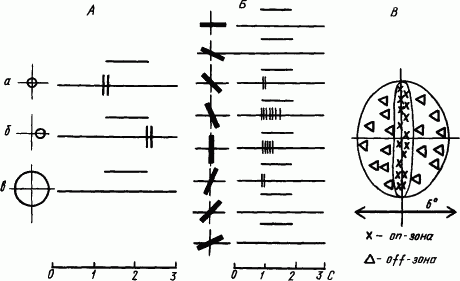
Рис. 54. Реакции нейронов зрительной коры мозга кошки на световое пятно (А, а, б), диффузное освещение (А, в) и световую полоску (Б) и общая форма вытянутого рецептивного поля (В):
слева от каждой нейрограммы указана форма стимула (кружок, полоска) и его локализация относительно рецептивного поля (крестик), над каждой нейрограммой чертой обозначена длительность стимуляции
Систематические исследования реакций нейронов зрительной коры мозга 17-го, 18-го и 19-го полей на световые стимулы разной формы и интенсивности, неподвижные и движущиеся, выявили важные закономерности формирования некоторых фрагментов нейрофизиологических механизмов зрительного восприятия (Д. Хьюбел, Т. Визел, 1974). Так, некоторые корковые нейроны наиболее активно реагировали на световую полоску, ориентированную вдоль вытянутого центра его рецептивного поля (рис. 54, Б). Как видно из рис. 54, наклоны полоски приводили к уменьшению ответа вплоть до его полного отсутствия. Такая избирательность коркового нейрона к ориентации линии находит объяснение в том, что на нем конвергируют сигналы от нескольких нейронов предыдущего этажа обработки зрительной информации (латерального коленчатого тела), и их концентрические рецептивные поля расположены в ряд, покрывающий световую полоску. Чем больше наклон полоски, тем меньше число вовлекаемых в реакции концентрических полей, а следовательно, и степень активации коркового нейрона. Если учесть, что в коре имеется множество нейронов с такими свойствами, но избирательно чувствительных к разной ориентации, то синтез их сигналов может дать описание всех видов линейных элементов контура воспринимаемого изображения. Такие нейроны называют простыми.
Другие нейроны зрительной коры имеют более сложную конфигурацию рецептивных полей, особенность которых состоит в отсутствии тормозного взаимодействия между участками поля. Поэтому они реагируют возбуждением на световой стимул, попадающий на разные его участки. Однако эти нейроны также строго избирательны к ориентации перепада освещенности. Вместе с тем их способность реагировать на стимул, последовательно проходящий ряд участков рецептивного поля, позволяет более точно оценивать его движение. При этом многие из них обнаруживают чувствительность лишь к одному направлению движения. Такие нейроны называют сложными.
Третий вид корковых нейронов, принимающих участие в обработке зрительной информации, характерен для рецептивных полей с комплексным возбудительным центром и тормозными зонами, ограничивающими размеры светового стимула, в частности длины его полоски. Эти клетки избирательно реагируют на ориентацию перепадов освещенности, линий и углов (главное, на их угловые размеры), оценивают направление движения стимулов. Такие нейроны называют сверхсложными.
Предполагают, что простые, сложные и сверхсложные клетки представляют три уровня синтеза результатов анализа зрительной информации. Свойства простой корковой клетки определяет конвергенция на ней сигналов от клеток латерального коленчатого тела, свойства сложной клетки — конвергенция на ней от простых клеток. Вопрос о происхождении сверхсложных клеток пока является дискуссионным.
Аналитико-синтетические процессы обеспечивают поэтапную обработку зрительной информации, направленную на восприятие целостного образа объекта. Изучение нейрофизиологических механизмов, осуществляющих эту обработку на разных этапах зрительного пути привело к представлению о четырех основных уровнях формирования целостного зрительного образа (В.Д. Глезер, 1985).
На первом уровне нейроны сетчатки и латерального коленчатого тела оценивают общую освещенность, выделяют сигналы из шума, подчеркивают контуры и дробят сетчаточное изображение на фрагменты.
На втором уровне нейроны 17-го поля коры определяют пространственно-частотные и ориентационные характеристики этих фрагментов. Здесь формируются группы нейронов, настроенные на разные ориентации и частоты, но направленные на один фрагмент. Они определяют его спектральный состав и связь с фрагментами иного спектрального состава.
На третьем уровне нейроны 19-го поля зрительной коры оценивают пространственное распределение возбуждения в нейронах предыдущего уровня и сопоставляют фрагменты изображения на основе их спектрального состава. Здесь также работают модули из нейронов разных свойств, обрабатывающих один участок зрительного поля, в результате чего выделяются отдельные компоненты фигур.
Наконец, четвертый уровень включает в себя обучающиеся нейроны, расположенные в нижневисочной и заднетеменной коре (НВК и ЗТК). Путем синтеза информации, поступающей от нервных механизмов предыдущего уровня, НВК обеспечивает опознание зрительного образа, ЗТК — его конкретизацию в пространстве.
Исследования процессов анализа и синтеза в процессах зрительного восприятия показали их значительную изменчивость в зависимости от многих условий: световой и темновой адаптации, контраста изображения с фоном, интенсивности светового стимула, внимания и т.п. На рис. 55 показаны изменения рецептивного поля коркового нейрона в разных условиях световой стимуляции. Эти и другие факты послужили основанием для гипотезы о том, что свойства нейронов зрительной коры определяются не столько жесткими отношениями конвергенции сигналов с предыдущих уровней, сколько адаптивной динамикой внутрикоркового взаимодействия нейронов, особенно тормозных.

Рис. 55. Изменения размеров рецептивного поля коркового нейрона при разном уровне зрительной адаптации (А), контраста светового стимула с фоном (Б) и энергии вспышки света (В) (по И.А. Шевелеву):
1, 2, 3 — реакция на вспышку (1 — 70 лк на фоне 0,1 лк, 2 — 0,7 лк на фоне 0,1 лк, 3 — 70 лк на фоне 10 лк), 4, 5 — реакция на световой квадрат поля (4 — 1 град2, 35 лк, 5 — 0,5 град2, 70 лк)
Изучения межнейронных отношений привели к заключению, что гибкая приспособительная изменчивость интегративных механизмов, в частности обработки зрительной информации, имеет в своей основе не жестко фиксированные связи между нейронами, а их вероятностное участие в формировании пространственно-временных мозаик возбуждающихся и тормозящихся нейронов. Мощное влияние на организацию аналитико-синтетической деятельности зрительной системы оказывают экологические различия образа жизни. Так, в зрительной коре у кошки преобладает ориентационная избирательность, у приматов — цветоразличение и стереоскопическое зрение, у белки — избирательность к быстрым движениям.
Многие принципы и способы организации нейрофизиологических механизмов аналитико-синтетических процессов зрительного восприятия оказываются общими и проявляются в деятельности всех сенсорных систем. Это относится прежде всего к механизмам оценки биологического значения действующих раздражителей. Такая оценка происходит на сравнительно высоком уровне интеграции поступившей сенсорной информации, ее сигнальной роли и текущего состояния организма. При этом мономодальные образы синтезируются в полимодальные и с учетом состояния организма, его потребностей и жизненного опыта формируется динамичный целостный образ воспринимаемого явления. В зависимости от того, является ли этот образ показателем благоприятных или опасных событий, возникает и соответствующая эмоциональная окраска восприятия.
Сложными процессами аналитико-синтетической деятельности мозга обусловлено возникновение новых видов приспособительного поведения животных. При этом формируются нервные механизмы объединения афферентных и эффекторных структур в целостную организацию достижения полезного результата, как это представлено в учении о функциональной системе (П.К. Анохин, 1968). Систематические исследования нейрофизиологического механизма формирования функциональной системы при осуществлении акта приспособительного поведения выявили определенную связь динамики вызванных потенциалов и нейронной активности с деятельностью ее основных блоков (афферентный синтез, принятие решения, программа действия, акцептор результатов действия).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
6.4. Аналитико-синтетическая деятельность коры больших полушарий
6.4. Аналитико-синтетическая деятельность коры больших полушарий Множество раздражителей внешнего мира и внутренней среды организма воспринимаются рецепторами и становятся источниками импульсов, которые поступают в кору больших полушарий. Здесь они анализируются,
9.4. Ассоциативные системы и интегративная деятельность мозга
9.4. Ассоциативные системы и интегративная деятельность мозга Даже простой выбор из нескольких инстинктивных программ требует «инстанции», где сравниваются значения сигналов внешней и внутренней среды организма. Точность правильного выбора, определяющего поведение,
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции Пер. А. НадирянВ этой и следующей главах дается краткое описание современного состояния эволюционной биологии, какой она была до 1995 года, когда возникло новое направление науки — сравнительная геномика.
8.2. Синтетическая теория эволюции
8.2. Синтетическая теория эволюции Популяционно-генетический подход заложил основы современной, так называемой синтетической теории эволюции (СТЭ), основанной на синтезе генетики и дарвинизма. Взаимосвязь между степенью генетической изменчивости популяции и скоростью
Глава 12 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЛИЧИНОЧНО-ХОРДОВЫХ КРУГЛОРОТЫХ И РЫБ
Глава 12 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЛИЧИНОЧНО-ХОРДОВЫХ КРУГЛОРОТЫХ И РЫБ Высшая нервная деятельность позвоночных животных отражает одну из важных тенденций их эволюции — индивидуальное совершенствование. Эта тенденция проявляется в нарастающей продолжительности
Глава 13 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЗЕМНОВОДНЫХ, ПРЕСМЫКАЮЩИХСЯ И ПТИЦ
Глава 13 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЗЕМНОВОДНЫХ, ПРЕСМЫКАЮЩИХСЯ И ПТИЦ Современные потомки первых обитателей суши сохранили в своей организации и поведении многие следы той ломки, которая сопровождала выход животных из водной стихии. Это видно, например, при
Глава 14 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ГРЫЗУНОВ И КОПЫТНЫХ
Глава 14 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ГРЫЗУНОВ И КОПЫТНЫХ После катастрофического конца эры холоднокровных гигантов, не сумевших приспособиться к новым условиям жизни, господствующее положение в животном мире заняли теплокровные млекопитающие. Высокий уровень обменных
Глава 15 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ХИЩНЫХ
Глава 15 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ХИЩНЫХ В жизни хищников приспособительное значение высшей нервной деятельности проявляется особенно ярко в жестокой борьбе за существование. Помимо непрерывной выработки все новых условных рефлексов защиты от более сильных врагов,
Глава 16 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ОБЕЗЬЯН
Глава 16 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ОБЕЗЬЯН Изучение высшей нервной деятельности обезьян представляет особый интерес по двум причинам. По-первых, обезьяны — самые высокоразвитые в психическом отношении животные, во-вторых, они самые близкие к человеку представители
Глава 17 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА
Глава 17 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА Жизнь на каждом шагу показывает неизмеримое превосходство разума людей над примитивными мыслительными способностями животных. Громадный разрыв между психической жизнью человека и животных долгое время служил поводом для
3.6. СИСТЕМНАЯ ДЕЯТЕЛЬНОСТЬ МОЗГА
3.6. СИСТЕМНАЯ ДЕЯТЕЛЬНОСТЬ МОЗГА Функциональное состояние центральной нервной системы непрерывно меняется в соответствии со сменой видов жизнедеятельности. Оно определяется позой, положением конечностей, множеством сигналов из внешней и внутренней сред. В каждый
НЕОДАРВИНИЗМ (СИНТЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ)
НЕОДАРВИНИЗМ (СИНТЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ) Если кого и можно назвать Ньютоном биологии, то, разумеется, Г. Менделя. Он сделал все, что требовалось для превращения биологии в «подлинную науку» наподобие классической физики, а именно:1) ввел невидимые сущности —
Глава 8 Пол мозга
Глава 8 Пол мозга No woman, no cry. Боб Марли У женщин проблема одна, Я повторяю всегда От Камчатки до Аргентины. Эта проблема — мужчины. Огден Нэш/Курт Вайль (пер. О. Волковой) Сосновая полевка (Microtus pinetorum) — моногамный вид грызунов: самцы помогают самкам выращивать потомство.
5. Анализаторная и синтетическая деятельность коры головного мозга
5. Анализаторная и синтетическая деятельность коры головного мозга Высшая нервная деятельность животного дает возможность выделять ему из сложной массы всевозможных раздражений, идущих из внешнего мира, те, которые имеют для животного то или иное значение. Собака при