Глава 9 ИЗМЕНЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА
Глава 9
ИЗМЕНЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ПРИ РАЗНЫХ СОСТОЯНИЯХ ОРГАНИЗМА
Гибкий и подвижный механизм высшей нервной деятельности обнаруживает громадные приспособительные возможности нормального функционирования при самых разных состояниях организма.
Знание наступающих при этом изменений нервных процессов помогает лучше понимать физиологическое действие указанных факторов и искать наиболее рациональные пути использования полезных и устранения вредных результатов таких воздействий.
Влияние физической и умственной работы
Влияние физической и умственной работы на процессы высшей нервной деятельности имеет большое практическое значение. Всякий труд, даже, на первый взгляд, чисто физический, связан со сложной высшей нервной деятельностью. Каждой профессии нужно учиться. Поэтому изучение влияния физической и умственной работы на условные рефлексы является ключом к решению основных проблем физиологии труда: тренировки, утомления, рациональной организации трудовых процессов и т.д.
Феномен Сеченова. Изучая закономерности утомления при мышечной работе, И.М. Сеченов обратил внимание на то, что лучшим видом отдыха является не бездеятельность, а смена деятельности с вовлечением в работу других мышечных групп. Так, если записывать при помощи эргографа ритмические поднятия груза одной рукой, то можно видеть, как постепенно высота поднятий уменьшается — развивается утомление (рис. 61, а). Перерыв в работе ведет к частичному восстановлению прежней высоты (рис. 61, б). Но если вместо отдыха поработать другой рукой (рис. 61, в), то восстановление оказывается гораздо более значительным (рис. 61, г).

Рис. 61. Влияние работы одной рукой на скорость восстановления работоспособности другой (по И.М. Сеченову):
а — запись работы правой рукой в течение 25 мин, б — продолжение записи после 10 мин отдыха, в — работа левой рукой в течение 2,5 мин, г — возобновление работы правой рукой
Эти явления объясняются следующим образом: когда в результате работы определенных мышечных групп наступает утомление (т.е. падение работоспособности), то это означает, что соответствующие клетки двигательного анализатора не в состоянии более поддерживать свое возбуждение и начинают переходить в тормозное состояние. Если теперь привести в действие другие мышечные группы, то среди корковых клеток двигательного анализатора возникнут новые очаги возбуждения. По правилу отрицательной индукции они будут оказывать тормозящее действие на ранее существовавшие очаги возбуждения и растормаживать возникшие очаги торможения. Такому растормаживанию и подвергнутся корковые клетки, которые были заторможены в связи с развивающимся утомлением.
Итак, утомление, развившееся при работе одних мышц, устраняется работой других мышц лучше, чем отсутствием всякой работы. В основе такого активного отдыха лежит явление отрицательной индукции в двигательном анализаторе с растормаживанием корковых клеток, оказавшихся в тормозном состоянии.
Феномен Сеченова послужил отправным пунктом для многочисленных исследований в области физиологии труда и спорта. С учетом благотворного влияния чередующейся мышечной деятельности строятся режим работы на производстве и тренировка спортсменов. На принципе активного отдыха основаны приемы производственной гимнастики, способствующей борьбе с утомлением. Тонизирующее действие мышечных упражнений человек испытывает каждый день, когда при помощи утренней зарядки рассеивает остатки сонного торможения и поднимает работоспособность своего мозга на более высокий уровень.
Мышечная работа. Феномен Сеченова представляет собой случай благотворного влияния умеренной мышечной работы на корковую деятельность. Однако с увеличением тяжести мышечной работы ее действие на процессы высшей нервной деятельности становится все менее благоприятным. Умеренная работа приводит к увеличению положительных условных рефлексов. Особенно возрастают условные рефлексы на сигналы, поступающие с кожного анализатора. Возможно, что в этом проявляется тесная связь кожного и двигательного анализаторов, обусловливающая их взаимодействие такого же типа, как взаимодействие между частями двигательного анализатора в феномене Сеченова. Что касается отрицательных условных рефлексов, то легкая работа, повышая возбудительные процессы, ведет к растормаживанию дифференцировок.
Однако при увеличении тяжести работы или увеличении ее продолжительности изменения процессов высшей нервной деятельности приобретают иной, прямо противоположный характер. Эти изменения заключаются прежде всего в том, что положительные условные рефлексы резко снижаются. В то же время дифференцировки становятся нулевыми. Видимо, ослабление возбудительных процессов приводит к относительному преобладанию тормозных. Выражением такого преобладания является также наблюдавшееся в этих опытах углубление последовательного торможения от дифференцировок.
Общие закономерности изменений высшей нервной деятельности в зависимости от тяжести работы подтвердились и в наблюдениях, проведенных на людях.
У спортсменов-подростков после забегов на разные дистанции отмечали затруднения в дифференцировании условных раздражителей и переделках их сигнального значения. Степень этих затруднений увеличивалась с тяжестью нагрузки.
Изменения условно-рефлекторной деятельности, наступающие при мышечной работе, зависят не только от тяжести работы, но и от ее характера. Особенно утомительна статическая работа, создающая длительное локальное возбуждение. Имеют большое значение и типологические различия. Так, например, в опытах с собакой сильного типа можно наблюдать увеличение условных рефлексов даже при очень тяжелой работе, при которой у собак с меньшей силой нервных процессов условные рефлексы уже резко снизили свою величину.
Таким образом, легкая мышечная работа, вызывая относительное усиление возбудительных процессов, увеличивает положительные условные рефлексы и растормаживает дифференцировки. Тяжелая мышечная работа, особенно статическая, перенапрягая возбудительный процесс в корковых клетках, приводит к падению положительных рефлексов и появлению охранительного торможения, т.е. дает картину утомления.
Умственная работа. Сведения о влиянии умственной работы на свойства процессов высшей нервной деятельности получены главным образом при исследовании условных рефлексов у учащихся или студентов.
Влияние умственной работы обычного учебного дня на различные условные рефлексы изучали, например, на группе студентов, у которых предварительно вырабатывали разнообразные условные рефлексы на простые звуковые и световые сигналы, а также связанные с использованием слова как специального раздражителя человека. К этому добавляли так называемый словесный эксперимент, в котором испытуемые отдавали отчет о своих реакциях во время хода опыта.
В результате таких исследований было выяснено, что после обычного 6-часового учебного дня наряду с некоторым нарастанием величины условных рефлексов на простые сигналы (например, звуковые) наступало удлинение латентного периода и снижение величины наиболее сложных условных рефлексов, осуществляемых с участием словесных раздражителей. В словесном отчете испытуемых появлялись ошибки. Так, например, испытуемый утверждал, что в процессе опыта он нажимал на рычаг только при слове «нажмите», в действительности же он производил нажатие и на простой звуковой сигнал. Заслуживает большого внимания тот факт, что в те дни, когда были занятия по физкультуре, изменения со стороны словесных условных рефлексов были выражены значительно слабее. Мышечная работа снимала утомление от умственной.
Описаны характерные изменения ЭЭГ при кратковременных умственных нагрузках и при разных видах умственной деятельности. Умственная работа сказывается и на вегетативных условных рефлексах. Наблюдения, проведенные на студентах, у которых были ранее выработаны сосудистые условные рефлексы, образованные на подкрепление охлаждением или нагреванием руки, показали, что после напряженной умственной работы, например во время подготовки к зачетам, эти рефлексы претерпевали резкие изменения. В большинстве случаев величина положительных условных сосудистых рефлексов становилась меньше, а некоторые полностью исчезали. Глубокие изменения наступали при этом в отрицательных условных рефлексах. Дифференцировочный раздражитель, дававший ранее только нулевые реакции, теперь стал вызывать резкое расширение сосудов. Таким образом, утомительная умственная работа может обусловить не только изменения величины условных рефлексов, но и их извращения.
Итак, умственная деятельность, т.е. специальная высшая нервная деятельность человека, вначале сама себя усиливает, но по мере своего продолжения начинает подвергаться ослаблению за счет развития тормозных процессов. В первую очередь страдают наиболее сложные специально «человеческие» условные рефлексы, связанные со словесными раздражениями. Чередование умственных занятий с физическими замедляет падение работоспособности корковых клеток.
Значение питания
На высшую нервную деятельность определенное влияние оказывает как общий недостаток или избыток пищи, так и недостаток или избыток отдельных ее составных частей — белков, углеводов, жиров, витаминов и неорганических веществ.
Общее голодание. Сытная еда снижает возбудимость нервных клеток пищевого центра. Испытанные непосредственно после еды пищевые рефлексы оказываются резко сниженными. Наоборот, задержка в поступлении пищи ведет к повышению пищевой возбудимости. Здесь имеет большое значение тип нервной системы. Поэтому условные пищевые рефлексы, например слюнные, у собак сильного типа оказываются тем выше, чем больший срок прошел после кормления. У собак слабого типа повышение пищевой возбудимости может быстро привести к достижению предела работоспособности корковых клеток. В таких случаях постановка опыта даже на 3–4 ч позднее обычного нередко ведет к снижению величины условных рефлексов.
Еще более резко выявляются изменения в условно-рефлекторной деятельности при длительном лишении пищи. Так, при суточном голодании происходят колебания в величине условных слюнных рефлексов, проявляющие фазный характер в зависимости от силы условных раздражителей: условные рефлексы на сильный звуковой сигнал в день голодания снижались, а условные рефлексы на слабые световой и кожно-механические сигналы повышались. В этом обнаруживается своеобразное действие голодания. Задержка в поступлении пищи приводит к повышению возбудимости пищевого центра, деятельность которого создает фон постоянного возбуждения в коре мозга. Поэтому слабые сигналы, вызывая ранее лишь небольшое условное возбуждение, теперь приводят к более значительному суммированию с фоновым возбуждением, и величина условных рефлексов на эти сигналы возрастала. В то же время сильные сигналы, вызывавшие и без того значительное условное возбуждение, при повышенной голодом возбудимости корковых клеток стали способствовать возникновению в них возбуждения, превышающего предел их работоспособности, так что рефлексы снизились (возможно даже запредельное торможение).
Показано, что изменения высшей нервной деятельности развиваются при голодании в следующем порядке.
1. Начальное повышение возбудимости коры с усилением условных рефлексов. 2. Ограничение нормальной условно-рефлекторной деятельности охранительным торможением. 3. Истощение процессов условного торможения с разрушением дифференцировок, запаздывания и т.п. 4. Истощение процессов условного возбуждения с исчезновением положительных условных рефлексов, вначале искусственных, затем и натуральных.
Белки, углеводы и жиры. Не только голодание, но и изменения в составе пищи влияют на состояние процессов высшей нервной деятельности. Если у крыс, получающих разное количество белка, вырабатывать оборонительные условные рефлексы на звуковые и световые сигналы, сопровождаемые электрокожным раздражением лапы, то прочность вырабатываемых рефлексов у них оказывается различной. Из рис. 62 видно, что у крыс, получавших наибольшее количество белков, условные оборонительные рефлексы оказались в среднем вдвое более прочными, чем у остальных. Еще более наглядно выступает влияние белка пищи на условные рефлексы крыс, если наблюдать за характером условных двигательных реакций. У крыс, находящихся в условиях белкового голодания, двигательная реакция на сигнал большей частью представляет собой очень слабое одиночное вздрагивание. В то же время двигательная реакция крыс, получавших много белка, состоит, как правило, из повторных рывков, не уступающих по интенсивности безусловной реакции на электрический ток.
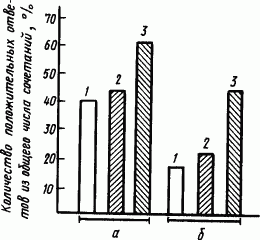
Рис. 62. Прочность оборонительного условного рефлекса на метроном у крыс при разном содержании белка в пище (по А.И. Макарычеву и М.А. Сергеевой): а — условная реакция отдергивания лапки, б — условная реакция писка; 1 — крысы на рационе, содержащем лишь следы белка, 2 — крысы на рационе, содержащем 18 % белка, 3 — крысы на рационе, содержащем 36 % белка
Если увеличивать или уменьшать порцию мяса в суточном рационе собаки (при неизменной его калорийности), то это заметно отражается на условных слюнных рефлексах. С первых же дней недостаток белка (0,2 г/кг массы животного) ведет к постепенному растормаживанию дифференцировок. Следовательно, наиболее уязвимыми в этом отношении оказываются процессы внутреннего торможения. Однако положительные условные рефлексы не обнаруживают значительных изменений даже при месячном содержании собаки на таком недостаточном белковом рационе.
Избыток белка (6,0 г белка/кг массы), наоборот, быстро улучшает дифференцировку, т.е. делает ее более полной. Положительные условные рефлексы возрастают по величине и становятся более прочными. Однако после содержания собаки примерно в течение 1 месяца на рационе питания с избытком белка наступают изменения другого рода. Дифференцировки остаются полными, но положительные рефлексы снижаются.
Существуют наблюдения, что повышенное содержание жира в пище собак ведет к изменениям условных слюноотделительных рефлексов, что свидетельствует о повышении возбудимости нервных клеток коры. Преимущественно углеводная пища вызывает противоположные изменения условно-рефлекторной деятельности, т.е. снижение корковой возбудимости.
Таким образом, недостаток белка сравнительно быстро приводит к ослаблению условного торможения, избыток — к его усилению наряду с усилением условного возбуждения. Длительное пребывание на избыточном белковом питании может привести к ослаблению возбудительных процессов. Преобладание жиров в пище способствует повышению возбудимости коры, а преобладание углеводов — ее снижению.
Витамины. Влияние авитаминоза C на высшую нервную деятельность изучали на обезьянах с выработанными условными пищевыми рефлексами нажатия на рычаг, за которым следовало открытие кормушки (Н.Л. Норкина, 1951). После содержания на пищевом рационе, лишенном витамина С, обезьяны в ответ на условный сигнал уже не нажимали на рычаг с прежней быстротой и силой. Движения их становились неуверенными: они направлялись то к рычагу, то к кормушке. В общем поведении животных также наступили перемены — исчезла свойственная им обычно подвижность и живость ориентировочных реакций. Ко времени, когда в крови оставались лишь следы витамина С, движения по условному сигналу к рычагу резко замедлились, число незавершенных реакций возросло и участились случаи полного выпадения условных рефлексов. Когда же обезьяну перевели на обычное кормление и уровень витамина С в крови достиг нормы, условные пищевые рефлексы быстро восстановились, возобновилась подвижность и прежняя игривость в поведении.
Таким образом, недостаток в пище витамина С ведет преимущественно к ослаблению возбудительных процессов в коре головного мозга.
Авитаминоз В1, как известно, приводит к тяжелым нервным расстройствам как движений, так и чувствительности (болезнь бери-бери). Опыты на собаках показали, что развитие авитаминоза B1 сопровождается резкими изменениями в высшей нервной деятельности в результате нарушения уравновешенности и общего ослабления тормозного и возбудительного процессов.
В то время как недостаток витамина В1 оказывает столь значительное влияние на высшую нервную деятельность, его избыток переносится организмом без вредных последствий. Так, при даче собакам с пищей значительных доз витамина B1 (50 мг в течение 100 дней) не было обнаружено никаких изменений условных рефлексов. Лишь подкожное введение растворов витамина В1 в дозах 10–20 мг вызывало временную неустойчивость условных слюнных рефлексов, нарушение силовых отношений и растормаживание дифференцировок.
Итак, избыток витамина B1 не нарушает нормального течения нервных процессов, недостаток же его ведет к быстрому ослаблению и нарушению нормальных соотношений между ними.
Влияние витамина D заключается в ослаблении положительных условных рефлексов и продлении следов действия отрицательных. Например, если на фоне дачи витамина D испытывать действие положительных раздражителей вслед за дифференцировкой или условным тормозом, то положительные рефлексы оказываются полностью заторможенными. Поскольку при введении витамина D происходит повышение в крови содержания солей кальция, весьма вероятно, что последний может обусловливать наблюдаемые изменения высшей нервной деятельности. Это тем более вероятно, что гормоны паращитовидных желез, также повышающие уровень кальция в крови, действуют на условные рефлексы подобно избыточному введению в организм витамина D.
Таким образом, влияние витамина D на высшую нервную деятельность, видимо, связано с действием солей кальция и выражается в усилении тормозных процессов с относительным ослаблением возбудительных.
Гормональные влияния
Железы внутренней секреции являются исполнителями многих рефлекторных регуляций. Вместе с тем гормоны поддерживают нормальное функциональное состояние и самих нервных клеток. Поэтому при нарушении работы желез внутренней секреции, например в связи с возрастными или патологическими изменениями в организме, нарушается и нормальный ход высшей нервной деятельности.
Половые гормоны. Общеизвестно, что в определенные периоды жизни человека, например в годы полового созревания или к старости, изменяется и состояние его психики. В этом проявляются изменения гормональных функций половых желез.
Влияние половых гормонов на процессы высшей нервной деятельности изучали в разных аспектах. При половом возбуждении были отмечены неустойчивость и снижение величины ранее выработанных у собак условных рефлексов и трудности образования новых. Действие мужских половых гормонов на условные рефлексы выявили на собаках-самцах в опытах во время полового возбуждения, вызванного самкой в период течки. В этих условиях наблюдали сначала повышение всех ранее выработанных условных рефлексов, а затем их снижение с развитием уравнительной и парадоксальной гипнотических фаз. При повторном половом возбуждении его тормозное действие на условные рефлексы не угасало. На этом основании наблюдаемое торможение было отнесено к типу безусловного.
О влиянии женских половых гормонов на высшую нервную деятельность можно судить по резкому изменению условных рефлексов, которое наблюдали у самок в период течки, а также время беременности и лактации. Так, во время течки у кошек условные рефлексы уменьшались, латентный период удлинялся. Новые условные рефлексы образовывались с трудом и были крайне неустойчивы.
Резкие изменения условно-рефлекторной деятельности под влиянием половых гормонов происходили у обезьян. Так, у самцов низших обезьян в периоды усиленной половой активности затруднялось образование условных пищевых рефлексов и нарушались ранее выработанные вплоть до отказа от еды. У половозрелых шимпанзе спокойного флегматичного типа в стадии отека наружных половых органов, когда содержание половых гормонов в крови достигает максимума, условные и безусловные рефлексы возрастали и становились более регулярными, а у беспокойных легко возбудимых рефлексы нарушались и часто выпадали. В период спаривания у самца гамадрилла, изолированного от самки, условный пищевой рефлекс не образовывался даже после 60 сочетаний, тогда как у самки он был выработан, как обычно, после 8 и упрочился после 20 сочетаний.
Общее значение постоянного влияния гормонов для процессов высшей нервной деятельности было выявлено в систематических и разнообразных исследованиях на кастрированных животных, проводившихся на протяжении многих лет в лабораториях И.П. Павлова. Оказалось, что у собак разного возраста и разного типа нервной системы в результате кастрации проявляются самые разнообразные отклонения в ходе условных рефлексов. Однако общим для всех кастрированных животных было резкое ослабление как возбудительного, так особенно и тормозного процессов. У всех животных, кроме того, появлялась так называемая циркулярность корковой деятельности, т.е. чередование периодов, отличающихся то высокими, то низкими величинами условных рефлексов. Наконец, почти у всех них развивалась сонливость. Таким образом, кастрация приводила к «инвалидности» нервной деятельности.
Резкое ослабление корковых процессов возбуждения и торможения после кастрации приводило также к тому, что любое перенапряжение этих процессов легко могло вызвать развитие патологических состояний высшей нервной деятельности по типу неврозов. Отмечают, что у самок кастрация вызывает менее значительные нарушения высшей нервной деятельности. Однако у собаки-самки слабого типа кастрация вызвала хаотичность условно-рефлекторной деятельности.
В действии половых гормонов на условные рефлексы обнаружены видовые различия. Так, кастрация вызывала резкое ослабление корковых процессов, особенно тормозных, у обезьян и лошадей, но сравнительно мало отражалась на условно-рефлекторной деятельности котов и кроликов.
Изменения высшей нервной деятельности при кастрации очень сходны с изменениями, наблюдавшимися при старении, когда также затрудняется образование новых условных рефлексов. Однако ранее выработанные рефлексы оказываются более устойчивыми. У старых собак наблюдалась слабость внутреннего торможения. При этом растормаживались сначала тонкие дифференцировки, затем более грубые. Нарушения подвижности нервных процессов ярко проявились в опытах с двусторонней переделкой положительного и отрицательного условных рефлексов.
Известны попытки восстановления старчески измененной высшей нервной деятельности, как и других функций организма, при помощи половых гормонов. Так, в медицинской литературе описаны способы «омоложения» путем впрыскивания экстрактов половых желез (Е. Броун-Секар, 1889), перевязки семявыносящих протоков, пересадки половых желез. Однако точные наблюдения показали, что операции «омоложения» приводили лишь к временному улучшению состояния, после чего быстро развивались явления старческого упадка.
В случае беременности подопытных собак отмечали неустойчивость и снижение величины условных рефлексов, растормаживание дифференцировок. Исследования динамики состояния высшей нервной деятельности на протяжении беременности показали, что в ее 1-й период (20–22 дня) условные рефлексы становятся неустойчивыми с общей тенденцией к возрастанию их величины, во 2-й период (22–44 дня) рефлексы более устойчивы, но несколько уменьшаются, в 3-й — (44–60) вновь появляется неустойчивость условных рефлексов и в 4-й период (3–5 дней перед родами) происходит их резкое снижение. При этом во все периоды беременности безусловные секреторные рефлексы возрастали, безусловное торможение усиливалось, а условное ослабевало, что проявлялось в легком растормаживании дифференцировок.
Таким образом, можно заключить, что половые гормоны играют важную роль в обеспечении необходимой степени возбудимости и работоспособности корковых клеток.
Гормоны щитовидной железы. Гипер- или гипофункции щитовидной железы оказывают влияние на высшую нервную деятельность, что проявляется, например, в нарушениях психики человека на фоне заболеваний, вызванных изменением функции щитовидной железы.
При гипотиреозах (пониженная функция щитовидной железы) условные рефлексы вырабатываются с трудом, имеют большие латентные периоды, особенно замедлено образование речевых реакций, безусловные рефлексы снижены. Изучение условных пищевых рефлексов у детей при разной степени недостаточности функций щитовидной железы выявило слабую возбудимость и пониженную работоспособность коры больших полушарий головного мозга. Препараты тироксина способствовали в этих случаях увеличению условных рефлексов.
Удаление щитовидной железы у животных приводило к резкому падению возбудимости корковых клеток и ослаблению тормозного процесса. Условные слюнные рефлексы, особенно пищевые, вырабатывались с трудом. Дифференцировка удавалась лишь при оборонительном рефлексе с электрокожным подкреплением, и то при значительной силе тока.
У молодых животных после удаления щитовидной железы наблюдались особенно большие отклонения в выработке условных рефлексов по сравнению с контрольными животными одного и того же помета.
Не только недостаток гормонов щитовидной железы, но и избыток их (гиперфункция щитовидной железы) также приводит к нарушениям высшей нервной деятельности. Условные рефлексы при тиреотоксикозах вырабатываются быстро, но оказываются очень нестойкими, легко развиваются фазовые состояния, безусловные рефлексы резко увеличены.
В экспериментальных условиях собакам в пищу систематически добавляли высушенную ткань щитовидной железы, после чего вскоре наступало резко выраженное нарушение тормозных процессов. Собаки не только перестали отличать отрицательный условный раздражитель от положительного, но величина условного слюнного рефлекса на отрицательный сигнал иногда даже превышала величину положительного рефлекса. При дальнейшем кормлении собак щитовидной железой наступали резкие изменения и положительных рефлексов: латентные периоды их укоротились, а величина возросла. Все это свидетельствует о резком повышении возбудительного процесса в этот период.
В зависимости от дозы препараты гормона щитовидной железы могут различным образом влиять на корковую деятельность. Так, по показателям условных слюноотделительных рефлексов у собак тиреоидин в малых дозах усиливает и концентрирует возбудительные и тормозные процессы в коре мозга, а в больших дозах вызывает запредельное торможение.
По показателям условных двигательных пищевых и оборонительных рефлексов у кур и голубей тиреоидин в умеренных дозах повышает возбудимость больших полушарий, а в больших дозах и при длительном введении вызывает развитие явлений торможения.
Характер и степень нарушений условно-рефлекторной деятельности зависели от типа нервной системы подопытных животных. Так, у собак сильного типа тиреоидин вначале вызывал увеличиние условных рефлексов и растормаживание дифференцировок, а у собак слабого типа — сразу уменьшение как условных, так и безусловных рефлексов.
Следовательно, гормоны щитовидной железы оказывают специфическое действие на высшую нервную деятельность, ускоряя и усиливая протекание основных нервных процессов.
Влияние гормонов паращитовидных желез. Введение гормона паращитовидных желез вызывало у собак кратковременное повышение условно-рефлекторной деятельности, которое сменялось снижением величины условных рефлексов, ускорением их угашения, усилением последовательного торможения, развитием сонливости. При экстирпации паращитовидных желез наступает резкое ослабление и даже полное исчезновение условных и безусловных оборонительных рефлексов у собак. Кроме того, нарушаются отношения между возбудительными и тормозными процессами за счет ослабления возбудительного, создающего относительное преобладание тормозного.
Таким образом, гормон паращитовидной железы, видимо, путем регулирования уровня кальция в крови поддерживает работоспособность корковых клеток главным образом путем активации возбудительных процессов.
Гормоны надпочечников. Помимо фактов влияния патологии надпочечников на состояние психической деятельности человека, показано, что гормоны коркового слоя (кортикостероиды) снимают утомление, а гормон мозгового вещества (адреналин) активирует эмоциональную сферу.
Частичные экстирпации надпочечников у собак приводят к глубоким и длительным изменениям высшей нервной деятельности. Положительные условные рефлексы изменялись у животных разного типа нервной системы по-разному, более определенными были изменения отрицательных рефлексов.
Характерное нарушение тормозных процессов проявлялось в том, что угашение всех рефлексов протекало значительно медленнее, чем в норме, а восстановление угашенного рефлекса происходило быстрее. Кроме того, наступали резкие искажения силовых отношений раздражителей и реакций.
При удалении мозгового вещества надпочечников обнаружены разнообразные нарушения условных рефлексов, также зависящие от типов нервной системы подопытных собак. Но для всех подопытных животных характерным было ослабление процессов внутреннего торможения с 1-й недели после операции. Раздражительный же процесс сначала усиливался, а затем ослабевал. Эти нарушения были длительными и проявлялись больше у собак с малой силой корковых процессов. Безусловные слюнные рефлексы были повышены у всех животных с 9–10-го дня после операции и к 3-му месяцу превышали исходную величину примерно на 20 %.
Введение адреналина способствовало нормализации условно-рефлекторной деятельности. Однократное введение собакам кортизона в малой дозе вызывало значительное увеличение положительных условных слюнных рефлексов, сменявшееся их уменьшением (рис. 63).
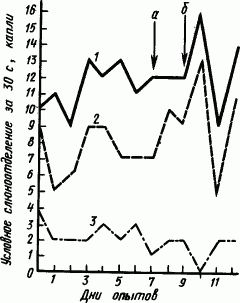
Рис. 63. Изменения условных рефлексов у собаки после введения кортизона в дозе 0,25 мг на 1 кг массы (по Н. Николаеву):
а — контрольное введение физиологического раствора, б — введение кортизона; 1 — слюноотделение на зуммер, 2 — то же, на свет, 3 — то же, на метроном 60 ударов/мин (дифференцировочный раздражитель)
Улучшалась и дифференцировка. Однако у животных со слабыми процессами возбуждения и торможения возникали нарушения условно-рефлекторной деятельности. Гидрокортизон, введенный крысам, укорачивал время поисков животными выхода из лабиринта и уменьшал количество ошибок в выборе ими пути.
Интересные данные были получены в опытах на собаках, которым вводился гормон гипофиза, усиливающий естественную выработку гормонов надпочечниками,— адренокортикотропный гормон (АКТГ). Наблюдение за слюнными условными рефлексами у этих животных показало, что характер влияния АКТГ очень зависит от типологических особенностей нервной системы собак.
Таким образом, гормоны мозгового и, в особенности, коркового вещества надпочечников могут усиливать и возбудительные, и тормозные процессы, повышая работоспособность клеток коры головного мозга и способствуя нормализации ее деятельности в случаях нарушения.
Гормоны гипофиза. Клиническая эндокринология дает много примеров того, что поражения гипофиза сопровождаются разнообразными нарушениями высшей нервной деятельности больных. Одни из влияний гипофиза на деятельность коры мозга могут осуществляться через стимулирующее действие гонадотропного гормона его передней доли на половые железы. По-видимому, таким путем некоторые формы гиперфункции гипофиза вызывают патологическое преобладание процессов возбуждения. Иное действие на процессы высшей нервной деятельности оказывают гормоны задней доли гипофиза. Так, введение больших доз этих гормонов собакам приводило к угнетению условных пищевых слюноотделительных рефлексов. Однако угнетение условных рефлексов наблюдали и в результате гипофизэктомии. Возможно, это объясняется относительным преобладанием в норме стимулирующего действия гонадотропного гормона.
Другим объяснением противоречивых результатов, полученных при изучении влияния гипофиза на условно-рефлекторную деятельность, может быть наличие фазности в его действии. Об этом свидетельствуют данные об изменении условных двигательных оборонительных рефлексов у собак под влиянием длительного применения препаратов гипофиза; показано, что это действие зависит от текущего состояния коры мозга.
Итак, гипофиз вторично повышает условно-рефлекторную деятельность с помощью гонадотропного гормона, а гормонами задней доли может ее стимулировать или угнетать в зависимости от количества гормона и длительности его действия.
Фармакологические воздействия
Изучение действия фармакологических веществ на высшую нервную деятельность животных и человека имеет значение для познания ее механизмов и восстановления ее нарушений. По поводу классификации веществ, действующих на высшую нервную деятельность, существуют разногласия, поэтому приводимая ниже классификация имеет условный характер.
Психотропные средства. В группе нейролептиков из средств адренолитического действия в психиатрии широко применяют аминазин как средство, снижающее возбуждение. В эксперименте на животных он обнаружил нормализующее влияние на условно-рефлекторную деятельность, нарушенную невротическим срывом. У собак-невротиков восстанавливались оборонительные условные рефлексы на ранее выработанный стереотип раздражителей. Однако величина положительных условных рефлексов под действием аминазина снижалась. Аналогичные результаты были получены и при изучении слюноотделительных условных рефлексов у собак, а также при исследовании условных рефлексов у обезьян. Аминазин оказывал угнетающее действие на двигательные оборонительные рефлексы у крыс, выработанные на электрокожном подкреплении. У кроликов аминазин в дозах 0,5–1,5 мк/кг снижал величину двигательных пищевых условных рефлексов.
Аминазин угнетает общее поведение, особенно ослабляет агрессивные реакции, нарушает дифференцировки и запаздывание. Однако в некоторых исследованиях отмечено первоначальное небольшое увеличение условных рефлексов. В зависимости от типа нервной системы собак одна и та же доза аминазина могла вызывать у них противоположные эффекты.
Под действием аминазина у кроликов становились реже ритмы электрической активности промежуточного мозга, увеличивалась амплитуда колебаний основных ритмов коры; активность альфа-ритма замедлялась до 4–5 колебаний/с; у людей усиливалась фоновая ритмика ЭКоГ.
При испытании действия разных доз аминазина на слюнные и двигательные рефлексы у собак оказалось, что в малых дозах он увеличивает латентные периоды и снижает величину положительных условных слюнных рефлексов, не нарушая дифференцировок, в то время как двигательные условные рефлексы осуществляются быстро и правильно, несмотря на сонливость животных. При увеличении дозировки нарастает угнетение всех условных рефлексов до полного их исчезновения.
Считают, что влияние аминазина на процессы высшей нервной деятельности связано с его избирательно угнетающим действием на восходящую активирующую систему ствола мозга. Однако показано, что он оказывает влияние и на многие другие структуры мозгового ствола. В частности, аминазин резко угнетает ядра заднего гипоталамуса, связанные с агрессивно-оборонительным поведением, и несколько повышает возбудимость гиппокампа.
Галоперидол также принадлежит к нейролептикам, но является производным бутирофенона. Он угнетает оборонительные условные рефлексы у крыс уже в очень малых дозах (0,1–0,2 мк/кг). Еще меньшие дозы нарушают процессы внутреннего торможения при дифференцировке (0,06 мг/кг) и угашении (0,02 мг/кг). Галоперидол — весьма эффективное средство при экспериментальных неврозах у кошек.
Резерпин (алкалоид из растения Rauwolfia) угнетает условно-рефлекторную деятельность, что показано на собаках, кошках, крысах, обезьянах. Он тормозит преимущественно оборонительные условные рефлексы, а также выработанные с условной реакцией страха.
Таким образом, нейролептические средства снимают избыточное возбуждение и могут выводить из невротических состояний, особенно связанных с дефицитом торможения.
В группу транквилизаторов входят такие вещества, как мепробамат, седуксен, элениум, диазепам.
Транквилизаторы в отличие от нейролептиков в обычных дозах, как правило, не угнетают классические пищевые и оборонительные условные рефлексы, а в малых дозах даже облегчают их, снимая напряжение тревоги и страха. Угнетение условных рефлексов большими дозами этих веществ при их повторных введениях исчезает, что объясняют адаптацией.
Транквилизаторы преимущественно действуют на лимбическую систему мозга человека, где происходит интеграция отрицательных эмоциональных реакций. Поэтому не образуются временные связи со словами, несущими отрицательные эмоции, при возможности их выработки на невербальные раздражители.
Таким образом, транквилизаторы, снимая состояние тревоги в малых дозах, улучшают условно-рефлекторную деятельность.
Антидепрессанты отличаются более широким спектром своего стимулирующего влияния, оказывая холинолитическое действие. Так, коразол оказывает «пробуждающее» действие при глубоком подавлении высшей нервной деятельности снотворными средствами или наркотиками. Стрихнин усиливает пищевые условные рефлексы, что было установлено еще в лаборатории И.П. Павлова. В дозе 0,088 мг/кг он увеличивал оборонительный условный рефлекс у кошек, а в дозе 0,2 мг/кг улучшал выработку дифференцировок у крыс. Пикротоксин (0,75–1,25 мг/кг) облегчает решение лабиринтных задач крысами.
Антидепрессанты активируют возбудительные процессы и способны восстанавливать высшую нервную деятельность, подавленную наркотиками.
Седативным средством усиления тормозных процессов является бром. Препараты брома, главным образом в виде бромистого натрия, издавна использовали в медицине в качестве успокаивающего средства. При этом действие брома расценивали как снижающее повышенную возбудимость нервной системы. Однако уже первые исследования изменений условных рефлексов после введения брома, произведенные в лаборатории И.П. Павлова, показали, что в основе благоприятного действия брома лежит не подавление процессов возбуждения, а усиление тормозных процессов. Выяснилось, что под влиянием введения брома усиливается выработка условного тормоза.
Например, у одной из очень возбудимых подопытных собак не удавалось образовать условно-тормозные комбинации из тона органной трубы и теплового сигнала условного пищевого рефлекса, несмотря на то, что эта комбинация применялась без подкрепления более 20 раз. После же введения брома звук трубы сразу приобрел способность тормозить условный рефлекс на тепло.
Введение брома улучшает также развитие запаздывания при отставленных условных рефлексах. Бром способствует концентрации тормозного процесса. Так, если у собаки в процессе опыта появляется сонливость, то ее можно устранять бромом.
При исследовании содержания брома в крови подопытных собак, у которых изучали тип высшей нервной деятельности, оказалось, что у собак сильного типа концентрация брома была выше (0,27–0,46 мг %), чем у собак слабого типа (0,12–0,29 мг %).
Описаны значительные видовые различия в чувствительности животных к брому. Так, бром не оказывает такого положительного влияния на процессы внутреннего торможения у голубей, как у собак, а при увеличении дозы быстро начинает их угнетать.
Усиливающее и концентрирующее тормозной процесс действие брома проявляется лишь при определенных его дозах, величина которых оказывается строго индивидуальной. Например, при изучении его влияния на условный отряхивательный рефлекс у кроликов оказалось, что дозы брома 0,1–1,0 г/кг усиливают дифференцировочное торможение, а 3,0 г/кг приводит к растормаживанию. При экспериментальном изучении типа высшей нервной деятельности бром используют как специальный индикатор силы тормозного процесса.
Препараты брома используют также для лечения нарушений высшей нервной деятельности, связанных с ослаблением тормозного процесса.
Таким образом, в определенных дозах седативные средства (в частности, бром) улучшают выработку всех разновидностей условного торможения, а также способствуют его концентрации. Большие их дозы ведут к перенапряжению торможения, в результате чего может произойти ослабление тормозного процесса.
Ноотропные вещества выделились недавно как средства, повышающие функциональное состояние высших отделов мозга и применяемые для лечения психических заболеваний. Наиболее типичный их представитель — пирацетам — в экспериментах ускорял выработку условных рефлексов избегания в лабиринте с водой у мышей, предотвращал у них выпадение памяти при гипоксии и оказывал многие другие положительные влияния на процессы высшей нервной деятельности. Пирацетам применяют при явлениях нарушения мозгового кровообращения.
Таким образом, ноотропные средства улучшают деятельность мозга, повышают работоспособность механизмов мышления.
Галлюциногены, как указывает их название, вызывают галлюцинации, преимущественно зрительные, появление фантастических образов, искаженного восприятия окружающего мира, ощущения собственного тела и времени. Так действуют индольные производные, из которых наиболее известен диэтиламид лизергиновой кислоты (ДЛК), производные некоторых катехоламинов (мескалин) и вещество, выделенное из гриба Amantia muscaria, а затем синтезированное (мускимол).
Все галлюциногены угнетают условные рефлексы. Особенно чувствительны к ним люди и обезьяны, у которых доза ДЛК, вызывающая нарушения, в сотни раз меньше, чем у собак, и в тысячу раз меньше, чем у крыс.
ДЛК в дозе 0,08–0,125 мг/кг и мескалин в дозе 11–15 мг/кг полностью подавляли у крыс пищевые и мало влияли на оборонительные условные рефлексы. Вместе с тем ДЛК в дозе 0,15 мг/кг замедляла угашение условного оборонительного рефлекса и нарушала дифференцировки, а мескалин в дозе 25 мг/кг, наоборот, ускорял угашение.
Таким образом, галлюциногены вызывают извращения восприятий и тормозят положительные условные рефлексы, действуя в основном на подкорковом уровне анализа поступающей информации.
Психостимуляторы. Так называют вещества, в умеренных дозах улучшающие высшую нервную деятельность, особенно возбудительные процессы. Типичный их представитель — кофеин. Он поступает в организм в составе чая и кофе — напитков, которые с незапамятных времен известны своим возбуждающим действием.
Специальные исследования действия кофеина на высшую нервную деятельность показали, что этот алкалоид усиливает процессы возбуждения в коре головного мозга. Он задерживает угасание условных рефлексов, устраняет запаздывание отставленных условных рефлексов и ведет к растормаживанию дифференцировок.
Однако такое действие кофеина можно объяснить не только усилением процессов возбуждения, но и угнетением другого нервного процесса — торможения. Подобно брому, кофеин оказывает свое усиливающее действие лишь в определенных дозах.
Предельные дозы кофеина колеблются от 0,03 до 0,5 г/кг для разных животных. Например, у коров подкожное введение 5–20 мл 20 % раствора натриево-бензойной соли кофеина (0,36–1,44 г чистого кофеина) уже вызывало явления торможения условных рефлексов типа запредельного. У голубей кофеин в дозах 0,002–0,01 г/кг усиливал положительные условные рефлексы, а в дозах 0,003–0,1 г/кг ослаблял их.
Однако усиливающее рефлексы действие кофеина, по-видимому, не сводится к простому повышению возбудимости корковых клеток. Так, прямое измерение порогов раздражения коры у кошек через хронически вживленные электроды показало, что несмотря на введение кофеина в дозах, увеличивающих условные рефлексы, пороги прямого раздражения корковых клеток не снижались, а при введении больших доз (свыше 100 мг/кг) даже возрастали, что может объяснить активацию условных рефлексов его влиянием на подкорковые уровни их осуществления.
Другой представитель психостимуляторов — фенамин. Действие фенамина (аналогичный ему заграничный препарат — бензидрин) исследовали при употреблении этого препарата людьми, в опытах на обезьянах, собаках, крысах и кроликах. Во всех исследованиях выявлено характерное свойство фенамина — усиливать процессы возбуждения в коре головного мозга особенно при утомлении. При этом оказалось, что чувствительность к фенамину сильно колеблется.
В опытах на обезьянах показано, что малые дозы фенамина (0,1–0,5 мг/кг) ускоряют, а большие дозы (1 мг/кг) замедляют условные оборонительные реакции. Действие фенамина было более выражено у животных слабого типа нервной системы или находящихся в сонливом состоянии.
Как показали наблюдения на человеке, прием фенамина в малых дозах способствует устранению сонливости и утомления. При этом человек лучше и точнее производит как физическую, так и умственную работу. В основе такого подъема работоспособности лежит усиление фенамином коркового процесса возбуждения. Действие фенамина длится несколько часов, после чего усиление возбудительного процесса сменяется его угнетением.
Исследования нейронной активности структур коры и подкорковых структур мозга кролика при выработке условного оборонительного рефлекса на звук показали, что под действием фенамина число «обучающихся» нейронов увеличивается в коре и уменьшается в стриопаллидарных образованиях.
Психостимулятором является и эфедрин — алкалоид, который применяется в качестве тонизирующего и возбуждающего средства. По своей химической структуре он близок к адреналину. Подобно адреналину, эфедрин оказывает благотворное влияние на нарушенную нервную деятельность. Однако у здоровых собак однократное введение эфедрина в так называемых средних терапевтических дозах для человека (0,2–0,5 мг/кг) ведет к снижению условных слюноотделительных пищевых рефлексов, главным образом на слабые положительные раздражители без нарушения дифференцировок. Увеличение пищевых условных рефлексов наступало лишь при гораздо, меньших дозах (0,005–0,06 мг/кг). Оборонительные двигательные условные рефлексы при многократных инъекциях эфедрина в дозе 1 мг/кг усиливались, а при дозе 10 мг/кг угнетались.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
6.6. Типы высшей нервной деятельности
6.6. Типы высшей нервной деятельности Условно-рефлекторая деятельность зависит от индивидуальных свойств нервной системы. Индивидуальные свойства нервной системы обусловлены наследственными особенностями индивидуума и его жизненным опытом. Совокупность этих свойств
Глава 1 ВОЗНИКНОВЕНИЕ, СОДЕРЖАНИЕ И МЕТОДЫ ФИЗИОЛОГИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 1 ВОЗНИКНОВЕНИЕ, СОДЕРЖАНИЕ И МЕТОДЫ ФИЗИОЛОГИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ С незапамятных времен внимание человека привлекали проявления психической жизни. Откуда берутся мысли? Как понимать сознание, вмещающее весь мир и управляющее нашими поступками? Что
Глава 5 ДИНАМИКА ПРОЦЕССОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 5 ДИНАМИКА ПРОЦЕССОВ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Вся сложная и разнообразная деятельность высших отделов нервной системы построена на работе двух основных нервных процессов — возбуждения и торможения. Протекая в подвижных пространственных и временных
Глава 8 ТИПОЛОГИЯ И ГЕНЕТИКА ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Глава 8 ТИПОЛОГИЯ И ГЕНЕТИКА ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Нервная деятельность, как мышечная сила, рост, цвет глаз и прочие признаки, может быть индивидуально различной. Эти различия во многом зависят от наследственности. В свою очередь, наследственность формируется
ГЛАВА 3. УПРАВЛЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТЬЮ (ПОВЕДЕНИЕМ)
ГЛАВА 3. УПРАВЛЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТЬЮ (ПОВЕДЕНИЕМ) Поведение собаки, т.е. сложная совокупность ее взаимодействий с окружающей средой, привычно рассматривается человеком с позиций собственного душевного мира. Такой подход к психике животных хорошо объясним и,
3.10. НАРУШЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
3.10. НАРУШЕНИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ Сочетание нейрохирургических вмешательств в определенные отделы мозга и условно-рефлекторного метода привело к пониманию многих сторон высшей нервной деятельности. Анализ ее нарушений расширил представления о работе мозга в
ОБЪЕКТИВНОЕ ИЗУЧЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ [ 20 ]
ОБЪЕКТИВНОЕ ИЗУЧЕНИЕ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ[ 20 ] Второй раз я имею высокую честь и глубокое удовлетворение принимать участие в торжестве открытия деятельности чрезвычайных русских обществ, народившиеся здесь, в Москве, и свидетельствующих о том, как
ФИЗИОЛОГИЯ И ПСИХОЛОГИЯ ПРИ ИЗУЧЕНИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ [ 25 ]
ФИЗИОЛОГИЯ И ПСИХОЛОГИЯ ПРИ ИЗУЧЕНИИ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ[ 25 ] Прежде всего я считаю своим долгом благодарить Философское общество, что оно в лице своего председателя изъявило готовность выслушать мое сообщение. Мне трудно было сообразить, насколько это
ЭКСПЕРИМЕНТАЛЬНАЯ ПАТОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ [ 51 ]
ЭКСПЕРИМЕНТАЛЬНАЯ ПАТОЛОГИЯ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ[ 51 ] Несколько вступительных слов о сложной судьбе нашей работы по физиологии и патологии высшей нервной деятельности, принимая, что прилагательные «высшая нервная» отвечают прилагательному
ОБЩИЕ ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ И ЧЕЛОВЕКА1
ОБЩИЕ ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ ЖИВОТНЫХ И ЧЕЛОВЕКА1 Образы, картины поведения как нас самих, так и близких к нам высших животных, с которыми мы находимся в постоянных жизненных отношениях (как, например, собак), представляют огромное разнообразие, прямо
ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ В СВЯЗИ С НЕВРОЗАМИ И ПСИХОЗАМИ И ФИЗИОЛОГИЧЕСКИЙ МЕХАНИЗМ НЕВРОТИЧЕСКИХ И ПСИХОТИЧЕСКИХ СИМПТОМОВ [ 52 ]
ТИПЫ ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ В СВЯЗИ С НЕВРОЗАМИ И ПСИХОЗАМИ И ФИЗИОЛОГИЧЕСКИЙ МЕХАНИЗМ НЕВРОТИЧЕСКИХ И ПСИХОТИЧЕСКИХ СИМПТОМОВ[ 52 ] Из огромного материала, относящегося к изучению высшей нервной деятельности у собак по методу условных рефлексов, я остановлюсь
6. Типы высшей нервной деятельности
6. Типы высшей нервной деятельности На основании изучения условно-рефлекторной деятельности огромного числа собак академик И. П. Павлов создал свое учение о типах высшей нервной деятельности. Все многообразные особенности условно-рефлекторной деятельности собак
7. Патология высшей нервной деятельности
7. Патология высшей нервной деятельности Одной из сторон работ академика И. П. Павлова, имеющей большое значение для служебного собаководства, является развитие им учения о патологии нервной деятельности.Высшая нервная деятельность собаки может быть сорвана
Некоторые актуальные вопросы генетики поведения и высшей нервной деятельности[50]
Некоторые актуальные вопросы генетики поведения и высшей нервной деятельности[50] Исследования по генетике поведения начались почти с самого начала нынешнего столетия, когда законы Менделя стали достоянием широких кругов биологов.При этом выяснилось, что признаки