Глава 13 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЗЕМНОВОДНЫХ, ПРЕСМЫКАЮЩИХСЯ И ПТИЦ
Глава 13
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЗЕМНОВОДНЫХ, ПРЕСМЫКАЮЩИХСЯ И ПТИЦ
Современные потомки первых обитателей суши сохранили в своей организации и поведении многие следы той ломки, которая сопровождала выход животных из водной стихии. Это видно, например, при сопоставлении условно-рефлекторной деятельности земноводных пресмыкающихся. Экологические особенности жизни птиц в воздушной среде, определившие своеобразное строение, их мозга, нашли отражение в сочетании высокого развития у них как инстинктов, так и условных рефлексов.
Лягушки и жабы
Внимательное наблюдение за голосовыми реакциями лягушек в естественной обстановке позволяет уловить известный порядок. Лишь только одна лягушка начинает квакать, к ней сразу присоединяются остальные. Замолкают они внезапно все вместе. Подражая голосу лягушек, можно заставить их квакать. Это указывает на определенное биологическое значение голосовых сигналов, которые играют важную роль в половом поведении.
Другим важным звуковым сигналом для лягушки является «шлепание» при прыжке в воду. Если вспугнуть одну лягушку, то как только она прыгнет в воду, вслед за ней немедленно начинают прыгать и остальные. Это происходит даже, если поставить специальные перегородки, чтобы лягушки не видели друг друга. В этом случае, как и во многих других, трудно утверждать, является ли такая защитная реакция на звуковой сигнал натуральным условным рефлексом или она уже вошла в наследственную основу и стала врожденным инстинктом.
Ко всем другим звукам лягушка почти безразлична. Так, если она не видит человека, он может производить любые шумы — лягушка только настораживается. Но стоит человеку оказаться в поле зрения лягушки, как она обращается в бегство.
Своеобразны зрительные восприятия лягушек. Исследование электрических ответов сетчатки и зрительных долей мозга на освещение показало, что наибольшее возбуждение вызывают зеленые и синие лучи по сравнению с желтыми и красными. Вероятно, лягушки оказывают предпочтение зеленому цвету в результате обитания и поисков пищи среди зелени.
Сигнальным пищевым раздражителем для лягушек является не вид добычи, а ее движение. Так, вид бегающей мухи вызывает быстрое выбрасывание языка, которым лягушка ловит добычу. Однако неподвижную муху лягушка не схватит. Если приколоть муху на лавке перед лягушкой, то при каждом движении мухи лягушка бьет ее языком. Только после очень большого числа таких безрезультатных попыток лягушка отказывается от добычи. Натуральный пищевой рефлекс угасает. Однако достаточно сделать небольшой перерыв, и рефлекс восстанавливается.
Свойства искусственных условных оборонительных рефлексов у лягушек изучали, сочетая световые или звуковые сигналы с электрическими ударами через провода, положенные на полу. Такие рефлексы образовывались с трудом и оказывались непрочными. Некоторые исследователи даже ставят под сомнение возможность выработки таких рефлексов на световой сигнал. Однако возможность их выработки была подтверждена. Опыты показали, что лягушка не в состоянии образовать условный рефлекс защитного отдергивания только одной лапки. Причина, по-видимому, заключается и в еще малой двигательной специализации рефлексов, и в том, что для совершения прыжка лягушка всегда действует сразу обеими конечностями, поэтому ей несвойственно поджимание или отдергивание одной лапки, наблюдаемое у других животных.
Лягушка справляется только с самым простым лабиринтом. Поворот направо вел в тупик, где лягушку ударял электрический ток. Налево был свободный выход, через который она могла попасть в убежище — резервуар с водой. После 100–120 упражнений лягушка безошибочно выбирала правильный путь. Если стенки правой и левой сторон окрашивались разным цветом, то выработка реакции безошибочного выхода из лабиринта происходила быстрее. Если после усвоения правильного пути стенки перекрасить наоборот, то лягушка сбивалась с правильного пути. Этим демонстрируется роль зрительных сигналов в образовании условных рефлексов у лягушек.
Жаба может справиться с несколько более сложным лабиринтом и лучше различает цвета. Она может также отличить треугольник от четырехугольника, прямую линию от ломаной, но только когда они двигаются. Если к жабе поднести на ниточке вырывающуюся стрекозу, то она ее быстро схватит. Также схватит она и муху-ктырь. Попытается схватить и шмеля, но ужаленная им, отпустит его. Теперь, если поднести к ней муху-ктырь, похожую на шмеля, жаба ее не возьмет, хотя стрекоз она продолжает хватать.
Черепахи
Черепаха в случае опасности втягивает голову под панцирь и прикрывает ее передними лапами. Эту общую защитную реакцию оказалось возможным использовать для выработки искусственного условного оборонительного рефлекса. Его условными раздражителями можно сделать разные сигналы, в том числе и время. Такие условные рефлексы у болотной черепахи (Emys orbicularis) специализировались, угасали без подкрепления, растормаживались и снова укреплялись.
Локальные оборонительные условные рефлексы черепах изучали при подкреплении световых и звуковых, кожно-механических и щито-механических сигналов электрическим раздражением задней конечности, вызывающим разгибание передней. При отставлении безусловного подкрепления на 2–3 с такие условные рефлексы образуются на кожно-механические сигналы после <...> сочетаний, а на щито-механические — после 10–20 сочетаний. По мере укрепления величина условного рефлекса возрастала. При неподкреплении условный рефлекс быстро уменьшался и угасал полностью через 10–50 применений неподкрепляемого сигнала. Внешний тормоз (удар по щиту) временно снижал величину условного рефлекса.
Пищевые условные рефлексы у черепах, как и у рыб, изучали на основе пищедобывательных движений.
Таким движением являлось схватывание резинового мундштука, регистрируемое с помощью воздушной передачи (рис. 85). При каждом схватывании мундштука черепаху подкармливали. Если положительный сигнал, например желтый свет, при одновременном добавлении вертушки не подкреплялся, то после примерно десятка применений добавочный агент (вертушка) начинал обнаруживать свойства условного тормоза. Предшествование добавочного агента при дальнейшем совпадении с положительным затрудняло образование условного тормоза, но все же последний можно было выработать. Выработанный условный тормоз (вертушка) лишал действия и другие положительные сигналы (например, зеленый свет), к которым его присоединяли.
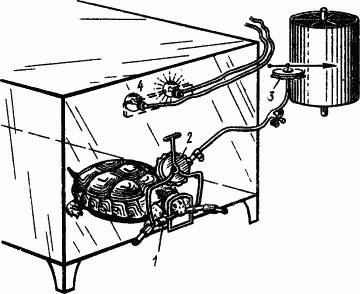
Рис. 85. Регистрация условных пищедобывательных рефлексов у черепахи (по В.В. Черномордикову):
1 — резиновый мундштук, который схватывает черепаха, 2 — воспринимающая капсула, 3 — регистрирующая капсула, 4 — лампочки для световых сигналов
Однако условный тормоз оказывался у черепах неустойчивым и для своего укрепления требовал до 100–150 применений. Оказалось даже, что при выработке условного тормоза с предшествованием добавочного агента часто вместо условного тормоза образовался условный рефлекс второго порядка. Попытки образовать цепной и следовой условные тормозы, применяя добавочный агент и положительный сигнал один за другим или с интервалом 5 с, оказались безуспешными. Следы раздражений в нервной системе черепахи непродолжительны. Не удалось также выработать условный тормоз из добавочного агента с иного анализатора (механического). Переделка сигнального значения условных раздражителей достигалась за 20–50 применений. Повторные переделки очень трудны и, как у рыб, приводили даже к ухудшению условно-рефлекторной деятельности.
Черепахи могут выработать реакцию безошибочного выхода из лабиринта с четырьмя тупиками. В первый раз черепаха добиралась до убежища (темное гнездо) в течение 35 мин, при второй пробе — за 15 мин, при третьей — за 5 мин. С каждым упражнением время прохождения лабиринта все уменьшалось и, наконец, при пятидесятой пробе черепаха добиралась до гнезда за 35 с. Описывают, что содержащиеся в террариумах черепахи выходили из гнезда при виде чашки, в которой им приносили воду для питья.
В опытах с обучением черепах у них удавалось вырабатывать зрительные дифференцировки: 1) черного и белого, 2) горизонтальной и вертикальной штриховки (если ширина штрихов была не менее 4 мм), 3) разной ширины штрихов (предельная разница 2–3 мм). Дифференцировки по яркости освещения вырабатывались с большим трудом, а по форме фигур (например, крест и звезда) совсем не получались.
Ящерицы, змеи и крокодилы
Легкий шорох немедленно вызывает стремительное бегство юркой ящерицы, даже когда она, казалось бы, дремлет с закрытыми глазами. Слуховые сигналы играют большую роль в ее жизни. В обстановке свободного поведения у ящерицы легко вырабатывается условный пищевой рефлекс подбегания к камню, на котором ее кормили при сочетании кормления с сигнальным звуком. Возможно, что резко выраженная реакция ящериц на звуки, напоминающие птичьи голоса, при полном безразличии к другим, даже более громким звукам, является натуральным оборонительным рефлексом. Показано значение гиппокампальной коры мозга ящерицы для образования условных рефлексов.
Немалую роль в поведении ящериц играет зрительный анализ. О наличии цветового зрения свидетельствует известная способность хамелеонов быстрым рефлексом хроматофоров изменять свою окраску соответственно цвету окружающего фона.
Ящерицы, как и змеи и крокодилы, хорошо ориентируются на местности в районе своего обитания. Некоторые змеи преследуют добычу, например мышей, по запаху следов. Уж отличает по запаху зеленую лягушку от жерлянки. После того как змее давали кусать железную палку, она некоторое время избегала всяких похожих на палку предметов.
Описывают, что крокодилы, живущие в зоопарке, приходили в сильное возбуждение, когда видели, что несут доску, на которой обычно разрезали для них мясо. О тонкости их зрительного анализа свидетельствует охота аллигаторов за прозрачными рачками, не видимыми человеческому глазу.
Птицы
В формировании поведения птиц большое значение приобретает жизненный опыт. Этому благоприятствует зрелость мозга птенца уже к моменту его вылупления. В добывании пищи и защите, в гнездостроении и выкармливании птенцов, во многих видах деятельности инстинкты птиц тесно переплетаются с разнообразными условными рефлексами, что приводит к формированию весьма сложных видов целесообразного поведения. Например, городские синицы (в ряде европейских стран) научились пробивать клювом крышки из тонкой фольги или бумаги и выпивать содержимое бутылок, оставляемых молочником у дверей хозяек. Описаны случаи, когда африканские стервятники приносили в клювах камни и разбивали ими добытые яйца страуса (И.И. Акимушкин, 1985).
Первые условные рефлексы у птенца. Еще находясь в яйце, цыплята инстинктивно реагируют на многие сигналы. Например, на слабый писк готовящегося выклюнуться цыпленка его братья и сестры немедленно откликаются из соседних яиц. Их энергичные движения заставляют яйца слегка ударяться друг о друга, усиливая общую активность. Громкий звук, например внезапный звонок, немедленно прекращает писк и движение. Цыплята в яйцах замирают в защитной неподвижности. Повторные звонки действуют все меньше и меньше. Сигнал опасности, не подкрепляемый действительным вредом, постепенно теряет свое значение, вырабатывается торможение этой сигнальной реакции.
Реакции цыплят, находящихся в яйцах, получили практическое применение для повышения продуктивности инкубаторного птицеводства. Так, искусственное воспроизведение акустических сигналов, даваемых курицей-наседкой, ускоряет и делает одновременным выклев цыплят, увеличивает привес при откорме, а фонетический анализ их ответного писка позволяет автоматически разделять петушков и курочек.
Вылупившийся из яйца цыпленок быстро осваивает двигательную координацию ходьбы и бега. К этому времени у него уже существуют основные безусловные рефлексы проглатывания попавшей в клюв пищи и движений защиты от повреждающих воздействий. Наряду с ними цыпленок обнаруживает врожденную инстинктивную реакцию клевания, для которой сигналом является вид мелких предметов. Следуя этому инстинкту, цыпленок клюет зерна, крошки, соринки, щепочки, пятнышки на полу, пальцы своих и чужих лапок и даже глаза других цыплят.
В природной обстановке такое клевание быстро приобретает условный характер, связываясь с видом клюющей наседки или петуха и звуками постукивания клювом. Поэтому ритмическое постукивание может поощрять склевывание корма, и цыпленок научается клевать более точно. С первых же часов жизни каждый цыпленок начинает обогащать фонд своей высшей нервной деятельности. Один, случайно склюнувший муху, становится ярым охотником за насекомыми. Другому на глаза попался червь, и он начинает рыться в земле.
Нескольким цыплятам были брошены мохнатые гусеницы Euchelia, которых взрослые птицы обычно избегают. Цыплята сразу их схватили, но тут же стали выплевывать и почесывать лапками клювики у основания — типичный для птиц «жест отвращения» при попадании в рот несъедобного. На другой день цыплята ходили вокруг предложенных им съедобных зеленых гусениц капустницы, но не брали их. Наконец, один из цыплят клюнул и проглотил гусеницу. Тогда и другие цыплята склевали остальных гусениц. Однако когда им снова предложили мохнатых гусениц, то цыплята остановились, почесывая клювики. Так они научились выбирать съедобную добычу.
Именно в первые дни жизни у птенцов формируются важнейшие условные рефлексы, которые определяют в дальнейшем их поведение. Так, цыплята, которые вскармливались только из рук человека, перестают реагировать на вид курицы и клохтание, которым она сзывает выводок. Вылупившиеся в инкубаторе птенцы фазана, куропатки, ржанки, водяной курочки и других бегали за человеком, который их кормил. Они не боялись постоянно находившейся в комнате собаки. Маленькие куропатки и водяная курочка клевали собаку, а трехдневный утенок пробовал жевать ее губы. Из этих наблюдений можно заключить, что их поведение в значительной мере определяется влиянием сложившихся в раннем возрасте условных натуральных рефлексов.
Уже в первые дни после вылупления птенцы мухоловки реагируют на прилет родителей пищевой реакцией, а на их «крики тревоги» — оборонительной реакцией затаивания. Регистрация ВП показала, что при этом в первом случае происходит усиление афферентации по зрительным путям, а во втором — их блокирование.
Свойства условных пищевых рефлексов у молодых цыплят изучали при помощи установки, позволяющей производить такое исследование одновременно у нескольких подопытных птиц (рис. 86). Оказалось, что уже в 6–8-дневном возрасте цыплята могут образовывать искусственные условные рефлексы высовывания головы в окно на звук звонка. Однако выработка дифференцировок по тону звонка (низкий сопровождался кормлением, высокий был дифференцировочный) явилась для цыплят трудной задачей. Они высовывали головки и на положительный, и на отрицательный сигнал. Мозг молодого цыпленка был еще не в состоянии ограничить иррадиацию возбудительного процесса рамками торможения. Только в 15–20-дневном возрасте у цыплят можно было выработать достаточно прочную специализацию условного рефлекса.
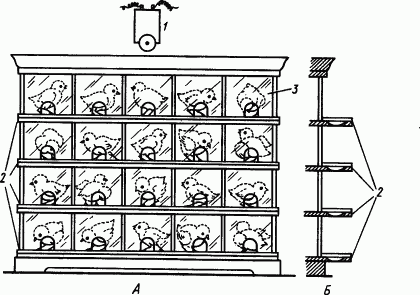
Рис. 86. Установка для одновременного исследования условных рефлексов в 20 цыплят. А, Б — вид спереди и сбоку (по Б.П. Фролову):
1 — звонок (условный раздражитель), 2 — щитки, открывающие сразу все кормушки, 3 — целлофановая передняя стенка с окошечками над кормушками
В условно-рефлекторной деятельности цыплят можно видеть значительные индивидуальные различия. Заслуживает большого внимания то обстоятельство, что цыплята, лучше вырабатывающие условные рефлексы, оказываются впереди других и по нагулу живой массы.
Условные рефлексы у голубей. Голуби, как и все птицы, выкармливающие птенцов, хорошо ориентируются на местности и легко находят свое гнездо. Они могут различать одинаковые на взгляд человека ящики и прямо подходят к тому, из которого получали корм. Отметив этот ящик, например, маленьким пятнышком, можно сильно облегчить птице задачу дифференцирования.
Вообще у голубя можно вырабатывать разнообразные условные положительные и тормозные рефлексы. Удавалось выработать условный тормоз, даже когда прибавочный агент подавали через иной анализатор (например, слуховой), чем условный положительный сигнал (зрительный). Удлинение паузы между прибавочным агентом и условным положительным сигналом могло приводить к образованию условного рефлекса второго порядка. Далеко отставленные и следовые условные рефлексы образовывались с трудом и оставались непрочными. После действия дифференцировочных раздражений была обнаружена положительная фаза индукции нервных процессов, а при действии гаснущих тормозов — отрицательная.
Пищевые условные рефлексы изучали у голубей в обстановке свободного поведения (И.С. Беритов, 1947). Сигналами служили световые фигуры в разных комбинациях.
Две кормушки для пищевого подкрепления располагали в противоположных концах камеры. Условный рефлекс подбегания к кормушке, расположенной под сигнальным транспарантом, образовывался после 4–5 сочетаний, был очень устойчивым с коротким латентным периодом (0–1 с). Подбегание к кормушке, расположенной напротив, вырабатывалось после 10–12 сочетаний, рефлекс был неустойчивым с более длительным латентным периодом 2–4 с. При выработке условных рефлексов на комплексные раздражители наблюдали широкую генерализацию рефлексов на все фигуры комплекса.
Выработка у голубей пищевых и оборонительных рефлексов на одновременные и суммарные комплексные раздражители, составленные из световых и звуковых сигналов, происходит лучше, чем на каждый из них в отдельности. Однако специализация рефлекса на комплекс и угашение реакций на его отдельные компоненты оказывается для голубя очень трудной задачей и наступает лишь после 150–200 применений. Попытки образовать у голубей временные связи на индифферентные раздражители без подкрепления пищевыми или оборонительными рефлексами («ассоциации») оказались безуспешными. Например, если выработать условный оборонительный рефлекс на вспыхивание лампочки, с которой предварительно неоднократно сочетался свисток, то сам свисток не вызывает рефлекса. Однако достаточно двух-трех подкреплений свистка оборонительной реакцией, и рефлекс вырабатывался. Следовательно, некоторое проторение пути «ассоциативной» связи в мозге голубя все-таки происходит.
У голубя четко проявляются вегетативные компоненты условных рефлексов. Например, после нескольких сочетаний звонка или метронома с вдыханием аммиака одно лишь применение звонка или метронома начинает вызывать резкое учащение дыхания и сердечных сокращений. Условный дыхательный рефлекс на зрительный сигнал (свет лампы), вырабатываемый при подкреплении вдыханием CO2 образуется после 3–8 сочетаний, а при подкреплении вдыханием NH3 после 1–5 сочетаний.
Обусловленное образом жизни птиц высокое развитие зрительного анализа у голубей находит практическое применение. Так, на одной фармацевтической фабрике голубя использовали в качестве контролера на конвейере, по которому изготовленные таблетки шли в упаковку. У него была выработана реакция клевать и сбрасывать бракованные таблетки по признаку их отличия от стандарта.
Голубиная почта. Голубиная почта известна издавна. Еще в древнем Египте жрецы с помощью голубей извещали о начале разлива Нила. Почтовый голубь доставил султану в Каир первое сообщение о вторжении крестоносцев. Голубиная депеша принесла миру, тревожно ожидающему исхода битвы при Ватерлоо, весть о поражении Наполеона. Осажденный прусской армией Париж поддерживал связь с внешним миром при помощи более 600 почтовых голубей, доставивших за время осады более 100 тыс. корреспонденций. Интересно, что для перехвата и уничтожения этих голубей осаждающие использовали специально для того тренированных ястребов. Первую голубиную почту в России организовал между Петербургом и Москвой в 1891 г. профессор Московского университета А.П. Богданов.
В первой мировой войне участвовало около миллиона связных голубей. Русский миноносец, лишенный возможности использовать радио, предупредил голубеграммой Севастополь о прорыве в Черное море немецкого крейсера. Широко применяли голубиную связь во второй мировой войне. Союзники поставили памятник голубке, доставившей донесение, благодаря которому был спасен экипаж подводной лодки, подбитой фашистским самолетом. Главное достоинство голубиной связи в том, что расположение отправителя остается неизвестным противнику, в то время как расположение станций радиосвязи легко определяется пеленгацией.
Подготовка связного голубя заключается в выработке у него главным образом зрительных условных рефлексов на все более далекие от голубятни ориентиры. Сначала голубя выпускают недалеко от голубятни. Сразу после прилета его ждет лакомый корм. Затем расстояние увеличивают. В течение первого года тренировки достигается дальность до 200 км радиуса безошибочного возвращения. Лишь на второй год обучения это расстояние возрастает до 300 км. Однако голуби возвращались домой и с более далеких расстояний. Считают, что связной голубь находит дорогу домой не только по зрительным сигналам. Об этом свидетельствуют способность голубя находить дорогу в тумане, облаках и ночью, и его обыкновение набирать кругами большую высоту (до 3–5 тыс. м), прежде чем устремиться в прямолинейный полет. С такой высоты зоркий глаз голубя может увидеть очертания знакомого ландшафта на расстоянии до 100 км.
Предполагают также, что голуби каким-то образом используют магнитное поле Земли для ориентирования в полете. Описаны случаи, когда голуби, оказавшиеся вблизи мощного радиопередатчика, сбивались с пути и беспомощно кружились на месте. Укрепленный на голове голубя магнит при солнечной погоде не оказывал на птицу никакого влияния, но в густых облаках сбивал его с правильного пути. Меняя полярность тока в спиралях Гельмгольца, укрепленных на голове голубя, изменяли направление его полета.
О существовании «компаса» в голове у птиц свидетельствует эксперимент с молодыми скворцами, впервые перелетающими из Скандинавии в Англию (юго-восточное направление). Когда их поймали по дороге, привезли в Швейцарию и там выпустили, то они, продолжая юго-восточное направление пути, залетели в Испанию, однако взрослые птицы, многократно совершавшие такие перелеты, оказавшись в Швейцарии, полетели в ином направлении и прибыли в Англию к месту назначения. Они научились накапливать и хранить ориентационную информацию для коррекции своего полета (Н. Тинберген. 1978).
Певчие и «говорящие» птицы. Китайская легенда гласит, что соловей научил людей музыке. Однако и сами пернатые певцы должны обучаться своему искусству. Здесь происходит сложное взаимодействие инстинктивных и подражательных реакций. Канарейку, например, можно научить соловьиным песням при помощи напетой соловьем магнитофонной ленты. Даже воробей, воспитанный среди канареек, в меру возможностей своего голоса пытается подражать пению приемных родителей. Если птенца с момента вылупления воспитывать среди певчих птиц другого вида, то он будет петь их песни. Однако стоит ему услыхать мелодию своего вида и сейчас же выработанные связи немедленно резко затормаживаются инстинктивной реакцией на естественный сигнал. Такая перестройка часто сопровождается бурной эмоциональной реакцией: взъерошенная птица мечется по клетке и долго не может успокоиться.
За 300 лет клеточного разведения и содержания канарейки утратили способность вить гнезда. Но если воспитать канарейку в клетке, где вместо обычных гладких жердочек поместить различные веточки с сучками и развилками, то в результате тренировки она быстро восстановит свою инстинктивную гнездостроительную реакцию.
Много условных компонентов выявляется в акте кормления птенцов.
Например, лесной конек, в гнездо которого посадили птенцов горихвостки, вскоре стал откликаться на их крик. Когда горихвостки были выкормлены, коньку подсадили птенцов серых славок. Новые голоса вначале вызывали лишь ориентировочную реакцию, но после нескольких кормлений, услышав зов своих новых питомцев, конек спешил к ним с кормом. В то же время собственных детей, выведенных в другом месте, голос которых ему не приходилось связывать с кормлением, лесной конек гнал и клевал.
Образование условного рефлекса кормления объясняет также, почему маленькие птицы выкармливают кукушонка, убивающего их детей. Это не «слепота инстинкта». Если подсадить кукушонка к не кормившей его птице, она улетит от него прочь.
Условные пищевые рефлексы подбегания к кормушке изучали в опытах с чижами и дубоносами. Такие рефлексы вырабатывались у них за 40–50 сочетаний, а у щеглов — за 70–80 сочетаний. После 2–3-месячного перерыва в работе для восстановления рефлекса требовалось около 20 сочетаний. Нетрудно было выработать дифференцировку по яркости светового сигнала. Однако на звуковые сигналы устойчивого условного рефлекса выработать не удавалось.
С канарейками, щеглами, синицами, снегирями и клестами проводили опыты в лабиринте с пищевым подкреплением. В этих опытах, например, снегирь при первой пробе искал выход 50 мин и прошел за это время 814 м, а после 40 упражнений находил выход за 1–2 мин. Ориентирами для птиц в лабиринте служили главным образом зрительные сигналы. Изменение условий освещения, приводящее к иному расположению света и теней, ослабляло вплоть до исчезновения всю выработку реакции выхода из лабиринта. Слабое освещение затрудняло поиски правильного пути, а темнота делала их безрезультатными. Вместе с тем смазывание стенок пахучими веществами, удлинение или укорочение коридоров не оказывало заметного влияния на поведение птиц в лабиринте. Следовательно, обоняние, осязание и мышечное чувство не имели существенного значения в поисках правильного пути. Рассказы о «говорящих» птицах часто грешат антропоморфизмом, т.е. птиц ошибочно наделяют человеческим сознанием. Так, попугаям (Psittaci), которые точно, вплоть до интонаций, могут имитировать голос человека, приписывают способность понимать смысл произносимых ими слов. Но тогда следовало бы ожидать, попугаи могут вести между собой и с людьми осмысленный разговор. Кроме того, пришлось бы допустить, что, например, пересмешники (Miminae), прекрасно воспроизводящие лай собаки, мычание коровы, ржание лошади, хрюканье свиньи, кукарекание петуха и т.д., понимают «языки» этих животных. Беспочвенность подобных предположений очевидна. Подражание птиц человеческому голосу не имеет ничего общего со словесным выражением мыслей у человека.
Хищные птицы. Орел, парящий под облаками, охватывает взором громадные просторы лесов и лугов, степей и гор и, заметив жертву, камнем падает на нее.
Мелкие соколы, охотящиеся за грызунами, например кобчик, могут с большой высоты заметить полевую мышь и безошибочно броситься на нее, увидеть одиночную стрекозу на расстоянии <...>00[7] м. Более крупных хищных птиц, таких, как кречет и сапсан, издавна использовали для охоты за дикими гусями, лебедями, цаплями.
Соколиная охота была широко распространена в Древней Руси, где искусство обучения боевых птиц стояло очень высоко. Сначала у сокола затормаживали натуральные, в основном зрительные, рефлексы, надевая на голову клобучок. Потом, удерживая сокола на привязи, у него вырабатывали ряд искусственных условных рефлексов схватывания добычи по сигналам посвистывания, прищелкивания и покрикивания. Постепенно круг усвоенных навыков расширялся, и обучение переносилось в поле. Хорошо подготовленный сокол летит на большой высоте за хозяином и бьет сверху вспугнутую дичь. В Средней Азии с помощью беркутов охотятся за лисицами, зайцами и даже волками. На Кавказе с помощью прирученных ястребов-перепелятников добывают перепелов на осеннем пролете.
Хищные птицы, даже питающиеся падалью, например коршуны или грифы-стервятники, в поисках пищи руководствуются в основном зрительными, отчасти слуховыми, но не обонятельными натуральными условными сигналами. Об этом говорят следующие опыты наблюдения.
В поле выставили чучело лани из хорошо просушенной кожи, набитое травой, и на него немедленно набросились грифы. В то же время труп свиньи, прикрытый сверху тростником, лежал несколько дней, стал разлагаться, но пролетавшие над ним грифы ничего не замечали. Один орнитолог приманивал рябчиков на пищик, имитирующий их голоса. На него налетел ястреб-перепелятник и ударил в голову, пытаясь схватить мнимого рябчика. Здесь хищник явно ориентировался только на слух.
Водоплавающие птицы. Домашние гуси и утки отличаются от диких сородичей не только особенностями телосложения, но и совершенно иным поведением. У них образовались новые инстинкты, искусственно созданные человеком. В то же время у диких видов сформировались другие натуральные реакции, определяемые условиями их жизни.
На примере поведения водоплавающих птиц наглядно проявляется значение для условно-рефлекторной деятельности принципа «биологической адекватности» раздражителей. Например, очень сильные, но не встречающиеся в естественных условиях звуки звонка почти не вызывали у дикого гуся двигательных реакций и изменений частоты сокращений сердца. Вместе с тем треск ломаемых щепок, напоминающий хруст, который производят, пробираясь через заросли (возможная опасность), заставлял селезня прятаться, замедляя ритм сердечных сокращений, а звуки плеска воды (приближение преследователя) или свист (бросающийся сверху пернатый хищник) приводили к бурной реакции бегства и резко учащали сердцебиение. Однако даже не действующий сам по себе звонок можно легко было превратить в сильно действующий сигнал, если каждое его 2-секундное применение сопровождать сильным безусловным раздражителем, например вдыханием паров аммиака. При этом после 3–5 сочетаний на звонок образуется условный оборонительный рефлекс с резким учащением частоты сердечных сокращений, который оказывается довольно прочным.
Натуральным сигналом половой реакции селезня является своеобразный крик самки. Притягательную силу этого рефлекса используют на охоте, когда диких самцов приманивают «подсадными» домашними утками или имитируют крик самки звуками манка. На основе таких натуральных рефлексов могут образовываться разнообразные двигательные и дыхательные условные рефлексы.
У диких и домашних (пекинских) уток способность к их выработке оказалась несколько различной. Например, условный рефлекс на метроном с частотой 200 ударов/мин образовался у дикой утки на 7-м сочетании, а у домашней — на 10-м. Дифференцировка метронома с частотой 90 ударов/мин была достигнута у дикой утки на 6-м его применении, а у домашней — лишь на 38-м. Следовательно, высшая нервная деятельность дикой утки может обеспечить более быстрые, чем у ее домашних сородичей, приспособительные реакции на меняющиеся условия жизненной обстановки.
Однако острота слухового анализа даже у дикой утки оказалась сравнительно небольшой — выработать полную дифференцировку звуков метрономов 200 и 152 ударов/мин она была не в состоянии (рис. 87). Переделывая охотничьи инстинкты некоторых водоплавающих птиц, их используют для рыбной ловли.
Так, рыбаки южных морей обучают бакланов ловить рыбу и приносить ее хозяину. Для этого баклану надевают шею кольцо, препятствующее проглатыванию пойманной рыбы. Кольцо снимают лишь тогда, когда он принесет рыбу в лодку. Вначале баклана держат на привязи. Когда вырабатывается досрочно прочное торможение рефлекса проглатывания добычи и укрепляется положительный пищевой рефлекс на хозяина, то его выпускают в свободный полет. Вся охота превращается для баклана в цепь сигнальных пищедобывательных актов, за которыми следует пищевое подкрепление, когда, закончив ловлю, хозяин снимет с его шеи кольцо и покормит.
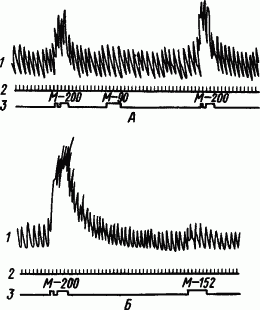
Рис. 87. Определение предела дифференцирования частоты стука метронома дикой уткой. А — полная дифференцировка; Б — неполная дифференцировка (по В.Г. Осиповой):
1 — дыхание, 2 — отметка времени, 3 — отметка сигналов
Куры. Домашние куры — прекрасная иллюстрация того, насколько глубоко переделывает человек природу животных в соответствии со своими потребностями. Мясные тяжелые кохинхинки, имеющие до 5 кг живой массы, не похожи на яйценосных легких леггорнов, дающих свыше 300 яиц в год, или бойцовых кур — участников азартных петушиных боев. Все они неузнаваемо отличаются по облику и поведению от своей прародительницы — дикой «курицы джунглей», которая несла 8–12 мелких яиц и весила 600–800 г.
Индийские веды упоминают о домашней курице около 2 тыс. лет до нашей эры. Однако, несмотря на столь длительный срок жизни под оценкой человека, куры сохранили высокое развитие условно-рефлекторных пищевых, защитных и половых приспособлений, очевидно, связанных со стадным свободным содержанием.
Домашняя курица образует разнообразные, по преимуществу пищевые и половые рефлексы. Все знают, как быстро привыкают куры ко времени и месту кормления и сбегаются со всех сторон на зов птичницы. Во время летнего выгула куры хорошо ориентируются в расположении своих полевых домиков. Известно также, как курица, высиживающая утиные яйца, приспосабливается к повадкам утят. Описывают, что курица, которая 3 года подряд выводила утят, сама водила последние выводки к ручью учиться плавать. Когда же на следующий год она вывела цыплят, то ее нашли в знакомом месте у ручья, куда она уже успела столкнуть клювом цыплят.
Как и все птицы, куры ориентируются главным образом с помощью зрения. В темноте курица не клюет, даже если стоит на куче зерен. Куры могут различать ящик, из которого получали корм, и пустой по рисункам на стенках. Они различали, например, три горизонтальные полосы и сплошное серое поле или черное пятно. Разницу в размерах они определяют лучше, чем в форме. Куры дифференцируют световые сигналы по интенсивности.
Если разложить в ряд рисовые зерна, причем одно класть свободно, а другое приклеивать так, чтобы его нельзя было склюнуть, курица быстро приучается клевать зерна через одно, даже когда они и не будут приклеены. Если приклеивать два зерна подряд и оставлять свободно лежащим третье, то курица приучается брать каждое третье зерно. Но если приклеивать подряд три зерна, оставляя свободным четвертое, то курица начинает путать. Результаты этих опытов иногда истолковывают, как доказательство умения курицы «считать» до трех.
Куры неплохо ориентируются по слуху. Например, когда на расстоянии 2 м друг от друга поставили две ширмы, за одной из которых находились пищавшие цыплята, то выпущенная за 15 м от ширм курица прямо бежала к той ширме, которая скрывала цыплят. После того как курице закрыли одно ухо, она стала ошибаться в определении места источника звука.
Свойства условного пищевого рефлекса высовывания головы в окно клетки на световые и звуковые сигналы изучали у петухов и кур. Рефлексы на метроном, звонок и вспыхивание лампочек оказались у петухов очень прочными, не угасали даже после 40 неподкреплений. Была образована дифференцировка метрономов 100 ударов/мин (положительный сигнал) и 40 ударов/мин. Удавалось также вырабатывать реакции движения к разным окнам на различные звуковые сигналы. После 6-месячного перерыва в работе в первый же опытный день все положительные условные рефлексы были восстановлены, дифференцировки пострадали в большей степени. Условные рефлексы у петухов оказались более устойчивыми, чем у кур.
На основе естественных пищедобывательных движений клевания, подкрепляемого кормлением, у кур вырабатывали условные пищевые рефлексы на звуковые и световые сигналы (рис. 88).
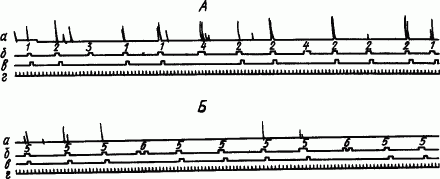
Рис. 88. Условное торможение пищедобывательных условных рефлексов у кур. А — совпадающий условный тормоз; Б — следовый тормоз (по А.В. Бару):
а — запись клевательных движений, б — отметки действия условных раздражителей, в — метки пищевых подкреплений, г — отметки времени (5 с); 1 — метроном, 2 — звонок, 3 — зеленый свет совпадает с применением метронома, 4 — белый свет в сочетании со звонком, 5 — зеленый свет, 6 — зуммер присоединения к зеленому свету
Условные клевания регистрирующего рычага по сигналам света лампы и стука метронома вырабатывались после 8–12 сочетаний. Вначале было много межсигнальных клеваний. В этом проявлялся генерализованный рефлекс на обстановку камеры. После 35–160 сочетаний рефлекс специализировался, и куры клевали рычаг только по сигналу. Полное угашение условного рефлекса наступало после 20–25 неподкреплений. Дифференцировка световых сигналов появилась после 5–7 подкреплений и стала прочной после 30 применений дифференцировочного раздражителя. Совпадающий условный тормоз образовался очень быстро и стал прочным после примерно 10-кратного применения тормозной комбинации. Цепной условный тормоз вырабатывался труднее и укрепился только после 70–90 применений. Еще труднее шло образование следовых условных тормозов с 5- и 10-секундной паузой между действиями тормозного агента и положительного условного раздражителя. При этом даже после 100 применений тормозной комбинации не удалось сделать условный тормоз прочным, а условное торможение концентрированным. После каждого применения такого тормоза развивалось сильное последовательное торможение. Выработка цепного и особенно следового условного тормоза проходила через фазу образования условного рефлекса 2-го порядка.
Биологическое значение сигналов условно-рефлекторной деятельности кур во многом зависит от их экологической адекватности. Так, дыхательные условные рефлексы на крик курицы имеют более короткий латентный период, бо?льшие величину и проявляемость, чем выработанные на тон генератора, сильнее выражена активность нейронов гиппокампа.
Высшая нервная деятельность и продуктивность домашних птиц. Птицеводы знают, как снижается яйценоскость кур в результате скученности. Например, в одном опыте уменьшение площади с 0,43 до 0,29 м2 в среднем на одну курицу привело к недополучению в среднем 19 яиц от каждой несушки. Одна из причин этого — торможение развития яйца оборонительными рефлексами, которые закрепляются в виде условных рефлексов на обстановку. Отсюда понятно, как важно сохранить спокойную обстановку в птичнике.
Биология птиц определила исключительное значение натуральных световых раздражителей в их поведении. Рассвет является сигналом для начала пищевой, половой и прочей деятельности, с наступлением темноты куры устраиваются на насестах и засыпают, чтобы заставить птиц больше кормиться и больше откладывать яиц, в практике птицеводства применяют так называемый продленный световой день.
Обычно включают дополнительное электрическое освещение раним утром и вечером и короткие осенние и зимние дни. При этом соответственно увеличивают дачу зерна в рационе. Иногда используют искусственное освещение для проведения дополнительного вечернего кормления. Разбуженные ярким светом куры поедают корм, затем свет тушится, и они вновь засыпают. Опыт многих птицеферм показал, что во избежание истощения курицы общее время ее светового дня не следует сразу увеличивать более чем до 12–14 ч. О хозяйственной эффективности такого использования натуральных световых сигналов свидетельствует сводная таблица результатов введения продленного светового дня на птицеферме (табл. 16).
Таблица 16. Влияние продленного светового дня на яйценоскость кур (по В.Ф. Ларионову)
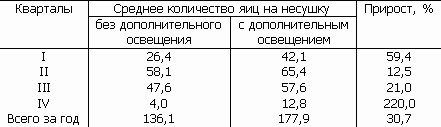
Большие практические перспективы открывают опыты по переделке суточного ритма сна и бодрствования у кур путем более частой смены света и темноты, приводящей к образованию условного ритма двух дней и двух ночей в одни календарные сутки.
В этих опытах группу цыплят с момента вылупления и группу взрослых кур помещали в птичник, в котором чередовали освещение и затемнение в течение каждых суток в следующем порядке: 0–4 ч — обычная ночь, 4–12 ч — день, 12–16 ч — «вторая ночь», 16–24 ч — «второй день». Птичник освещали лампами дневного света и обычными лампами, которые включали и выключали постепенно, имитируя рассвет и сумерки. Контролем служили группы цыплят и взрослых кур в условиях одного светового дня с 4 до 20 ч, равного общему времени освещения птиц подопытных групп.
Годичное содержание взрослых кур в условиях «двухдневных» суток привело к заметным изменениям в суточном распределении времени кладки яиц. Куры начали нести яйца в необычное для них время искусственного дня — с 20 до 24 ч. Резко увеличилась кладка яиц в его первую половину с 16 до 20 ч. Еще более заметные сдвиги по сравнению с контролем произошли у молодых несушек, воспитывавшихся в этих условиях с раннего возраста (рис. 89).
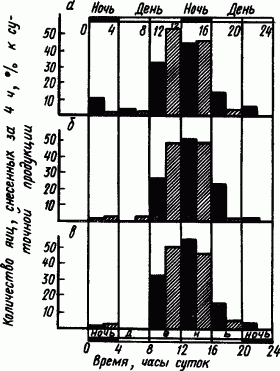
Рис. 89. Распределение кладки яиц в течение суток у молодых несушек, воспитанных в условиях обычных однодневных (штрихованные столбики) и «двухдневных» (черные столбики) суток (по М.Е. Лобашову, В.Б. Савватееву):
а — ноябрь — февраль, б — март — июнь, в — июль — октябрь
Выработка двухдневного суточного ритма оказала такое сильное действие на организм курицы, что у подопытных птиц втрое увеличилось число случаев, когда они несли яйца два раза в сутки. Образовались и укрепились новые условные рефлексы на время.
В середине искусственной ночи (с 12 до 16 ч) и перед искусственным рассветом раздавалось пение петухов. Проснувшиеся куры начинали обычную деятельность. Весь искусственный день почти до полуночи жизнь в подопытном птичнике кипит ключом. Петухи «ухаживали» за курами, дрались между собой, куры разгуливали по выгульным площадкам и рылись в траве, разыскивая корм. Когда однажды свет был погашен на 1 ч раньше, то кур не удавалось загнать на насесты. Они выскакивали через лаз наружу и разгуливали по птичнику.
Новый условный рефлекс на время оказался сильнее врожденной реакции на затемнение. Актографический контроль показал, что благодаря дневному сну общая суточная активность кур в условиях двухдневных суток выше, чем при непрерывном 16-часовом световом дне.
Перестройка жизненного ритма, усиление половой активности и питания, вызванные воспитанием в ритме двухдневных суток, имели результатом повышение яйценоскости на <...>0–30 % главным образом за счет зимних месяцев.
Двукратная смена дня и ночи в каждые астрономические сутки ускорила развитие и пришла к более раннему половому созреванию петушков. Половая активность последних была значительно более высокой, чем контрольных. Эти различия в поведении имели и морфологические основания в развитии половых желез (у контрольных петухов средняя масса семенников составляла 0,063 г, у подопытных того же возраста — 0,115 г, т.е. была почти вдвое больше).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Зимняя спячка земноводных
Зимняя спячка земноводных По образу жизни и устройству класс земноводных (Amphibia) является переходным между типично водными позвоночными (рыбы) и типично наземными животными. Известно, что различные виды лягушек, тритонов, саламандр и других, населяющих страны с умеренным
§ 43. Нервная система и органы чувств птиц
§ 43. Нервная система и органы чувств птиц Нервная система птиц состоит из центрального и периферического отделов. Головной мозг птиц крупнее, чем у любых современных представителей рептилий. Он заполняет полость черепа и имеет округлую форму при небольшой длине (см. рис.
Глава 10 Нервная система
Глава 10 Нервная система ГипнотизмДругая разновидность заболеваний, которые не подпадают под теорию Пастера, — это заболевания нервной системы. Такие заболевания смущали и пугали человечество испокон веков. Гиппократ подходил к ним рационалистично, однако большинство
Глава 7 АНАЛИТИКО-СИНТЕТИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ МОЗГА
Глава 7 АНАЛИТИКО-СИНТЕТИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ МОЗГА Вся высшая нервная деятельность состоит из непрерывного анализа — расчленения раздражителей окружающего мира на все более простые их элементы, и синтеза — обратного слияния этих элементов в целостное восприятие
Глава 12 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЛИЧИНОЧНО-ХОРДОВЫХ КРУГЛОРОТЫХ И РЫБ
Глава 12 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЛИЧИНОЧНО-ХОРДОВЫХ КРУГЛОРОТЫХ И РЫБ Высшая нервная деятельность позвоночных животных отражает одну из важных тенденций их эволюции — индивидуальное совершенствование. Эта тенденция проявляется в нарастающей продолжительности
Глава 14 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ГРЫЗУНОВ И КОПЫТНЫХ
Глава 14 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ГРЫЗУНОВ И КОПЫТНЫХ После катастрофического конца эры холоднокровных гигантов, не сумевших приспособиться к новым условиям жизни, господствующее положение в животном мире заняли теплокровные млекопитающие. Высокий уровень обменных
Глава 15 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ХИЩНЫХ
Глава 15 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ХИЩНЫХ В жизни хищников приспособительное значение высшей нервной деятельности проявляется особенно ярко в жестокой борьбе за существование. Помимо непрерывной выработки все новых условных рефлексов защиты от более сильных врагов,
Глава 16 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ОБЕЗЬЯН
Глава 16 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ОБЕЗЬЯН Изучение высшей нервной деятельности обезьян представляет особый интерес по двум причинам. По-первых, обезьяны — самые высокоразвитые в психическом отношении животные, во-вторых, они самые близкие к человеку представители
Глава 17 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА
Глава 17 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА Жизнь на каждом шагу показывает неизмеримое превосходство разума людей над примитивными мыслительными способностями животных. Громадный разрыв между психической жизнью человека и животных долгое время служил поводом для
ЭПОХА ЗЕМНОВОДНЫХ
ЭПОХА ЗЕМНОВОДНЫХ Земноводные были широко распространены и процветали в каменноугольный период (360–286 миллионов лет назад). Многие древние виды значительно превышали по размерам современные. Суровые, пустынные условия девонского периода (410–360 миллионов лет назад)
ЭПОХА ПРЕСМЫКАЮЩИХСЯ
ЭПОХА ПРЕСМЫКАЮЩИХСЯ Процесс эволюции некоторых древних земноводных в пресмыкающихся (рептилий) происходил в каменноугольный период (360–286 миллионов лет назад). К тому времени, как роскошные обширные болота — настоящий рай для земноводных — сменились унылыми пустынями
Глава XIII Нервная система
Глава XIII Нервная система Функции У нервной системы живых существ имеются две основные функции. Первая — сенсорное восприятие, благодаря которому мы воспринимаем и постигаем окружающий мир. По центростремительным чувствительным нервам импульсы от всех пяти органов
Высшая нервная деятельность
Высшая нервная деятельность За 20–25 дней до начала опытов была сделана попытка охарактеризовать основные особенности нервных процессов каждой подопытной собаки, для чего проводились обследования с помощью проб, подробно описанных на с. 90 этой книги. В силу