Глава 3 ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ПАМЯТИ
Глава 3
ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ ПАМЯТИ
Эффективность приспособительного поведения животных и успех разумной деятельности человека в значительной мере зависят от того, в какой мере животные используют свой жизненный опыт, а человек — полученные ранее знания для того, чтобы принимать правильные решения и избегать ошибочных. Эти знания извлекаются из памяти. Хотя в ее основе лежит нейрофизиологический механизм образования временных связей, понятие памяти имеет более широкий смысл. Рассмотренные в гл. 2 свойства и механизмы 0бразования условной связи раскрывают лишь одну начальную часть явлений памяти, объясняют, что означает запомнить. Другая, пожалуй, наиболее важная часть явлений памяти состоит в удержании на долгое время образовавшихся нервных связей, в способности помнить. Нередко под памятью подразумевают именно эту способность. Однако для того, чтобы воспользоваться полученной ранее полезной информацией, необходимо не только зафиксировать ее в новых нервных связях, не только сохранить эти связи, но и иметь возможность воспроизвести их в нужный момент, т.е. вспомнить.
Различия механизмов осуществления этих проявлений памяти наглядно демонстрируются врачебными наблюдениями за больными, у которых разные поражения мозга вызывали различные нарушения памяти. Например, описаны случаи, когда человек запоминает с трудом, но помнит долго или может запомнить лишь на очень короткое время или с большим трудом вспоминает то, что он легко запомнил.
Таким образом, понятие памяти включает в себя совокупность процессов фиксации, хранения и последующего считывания информации, получаемой организмом на протяжении его жизни.
Краткосрочная память
Давно замечено, что при некоторых заболеваниях мозга теряется способность запоминать события, происходящие во время болезни, хотя все бывшее ранее больной хорошо помнит. Эти явления в резкой форме были вначале обнаружены и описаны при тяжелой форме алкоголизма (С.С. Корсаков, 1880). Больной не был в состоянии запомнить происходящие события, но мог помнить, что происходило с ним раньше. Такую потерю памяти на недавние события наблюдали также у людей, переживших сотрясение мозга, при его очаговых поражениях, особенно в области височных долей, затрагивающих гиппокамп. Психологи отмечали сходные явления забывания событий, предшествовавших нервному потрясению. При этом память на все более давние события полностью сохранялась.
Такие явления ретроградной амнезии, т.е нарушения памяти на события, произошедшие незадолго до воздействия на мозг, при сохранности памяти на прежние события означают разные свойства памяти. На основании этих фактов сделано заключение о двух видах памяти: краткосрочной и долгосрочной (Д. Хебб, 1949).
Нарушения краткосрочной памяти типа ретроградной амнезии оказалось возможным воспроизвести в экспериментах на животных. Для этого достаточно было сразу после обучения подвергнуть мозг сильному воздействию и только что приобретенное знание исчезало, хотя ранее приобретенные навыки сохранялись. В экспериментах такими воздействиями были различные фармакологические средства, наркотики, переохлаждение мозга, нарушение его кислородного снабжения и другие средства. Однако чаще всего применяли электрическое раздражение мозга, вызывающее судорожные припадки.
На рис. 20 приведены результаты опыта, в котором у мышей вырабатывали условный оборонительный рефлекс избегания выхода на площадку, находящуюся под напряжением.
Получив удар током, мышь избегала выходить на эту площадку. Учитывали поведение мыши в первые 30 с после выхода на площадку. Как видно из рис. 20, если эфирный наркоз был дан сразу или на протяжении не более 10 мин после удара током на площадке, то условный рефлекс ее избегания был нарушен, мышь продолжала выбегать на площадку. Если наркоз был дан позже, спустя 16–20 мин, то он разрушал рефлекс не полностью, а наркотизация через 24 мин вообще не влияла на него.
Многие эксперименты показывали возможность избирательно выключать кратковременную память, не нарушая долговременную, и избирательно воздействовать на долгосрочную память, не нарушая краткосрочную. Такое действие оказывают, например, некоторые антибиотики, угнетающие синтез белка. Из этих данных был сделан вывод, что краткосрочная и долгосрочная память имеют разную природу и регулируются разными механизмами. Однако многие воздействия, выключающие краткосрочную память, как, например, нанесение на поверхность мозга раствора хлористого калия, нарушали и долговременную память. Вместе с тем описаны случаи, когда ретроградная амнезия через некоторое время проходила сама или снималась действием тока более слабого, чем применяемый в качестве подкрепления при выработке исследуемого рефлекса.
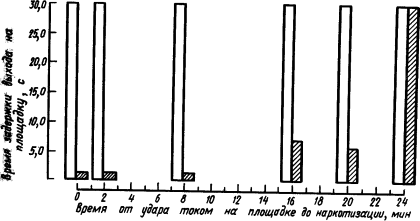
Рис. 20. Влияние наркоза на закрепление у мышей рефлексов избегания выхода на площадку, находящуюся под напряжением (по И. Абт и др.):
белыми столбиками изображено проявление рефлекса до наркотизации, заштрихованными — после наркотизации в разные сроки; остальные объяснения см. в тексте
На основании опытов с обезьянами было высказано предположение, что в условно-рефлекторной деятельности проявляются оба вида памяти: при отсроченных условных рефлексах — преимущественно краткосрочная, при следовых условных рефлексах — долгосрочная. Поэтому более вероятно, что механизмы краткосрочной и долгосрочной памяти тесно связаны между собой и являются последовательными этапами единого процесса, где памятный след, менее устойчивый на первом этапе, на втором этапе становится прочным. О природе двух видов памяти высказывают разные предположения, но, несмотря на интенсивные исследования, единой общепризнанной теории памяти пока не существует.
Возможные механизмы краткосрочной памяти. Наиболее распространено предположение, что краткосрочная память реализуется в виде многократной циркуляции потоков импульсов по замкнутым цепям нервных клеток. Основанием для такой гипотезы реверберации возбуждения были морфологические данные о наличии в сетях нейронов множественных соединений, образующих по петлям обратных связей круговые пути (Р. Лоренте де Но, 1934). Сочетания условного и безусловного раздражений активируют эти пути и многократное возбуждение нервных клеток, по которым протекают потоки импульсов, постепенно вызывает в них стойкие изменения, образующие долгосрочную память. Действительно, все воздействия, нарушающие краткосрочную память, прерывают возникающую циркуляцию потоков импульсов по замкнутым нервным путям.
Однако дальнейшие исследования показали, что в круговых путях нервных сетей находятся не только возбуждающие, но и тормозящие клетки, которые препятствуют «зацикливанию» циркуляции импульсов. На рис. 21 показана такая схема сети, в которой клетка 1 с клеткой 1А могла бы образовать круг реверберации импульсов, обусловливающей ритмическое возбуждение клетки Б. Но достижение порога возбуждения тормозящей клетки 1Б разрывает этот круг, затормозив клетку 1А.
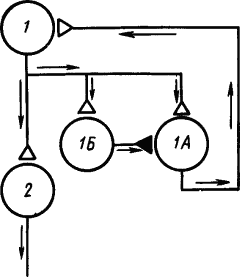
Рис. 21. Нервная цепь с замкнутыми нейронными цепями, по которым могут циркулировать потоки импульсов (по Э. Кенделу):
1 — корковый нейрон, получающий афферентные сигналы, 1А и 1Б — промежуточные нейроны, 2 — эфферентный нейрон; белыми треугольниками изображены возбуждающие синапсы, черными — тормозящий, стрелками показано направление движения потоков импульсов
Краткосрочная память не требует для своей реализации синтеза информационных (матричных) молекул РНК и ДНК. Она и не могла бы обеспечиваться этими биохимическими процессами как генетическая память, поскольку включение в полимер ДНК или РНК нуклеотидов происходит в сотни и тысячи раз медленнее, чем это необходимо для запечатления в них полной информации о текущих событиях. Вместе с тем при любом понимании ее природы, в том числе и реверберационном, речь идет о протекании потоков импульсов по новым для них путям, т.е. о временной синаптической проводимости.
Для объяснения механизма синаптической проводимости была предложена гипотеза о взаимодействии медиатора ацетилхолина с рецепторными белками постсинаптической мембраны и Na+, K+-АТФ-азой при разных концентрациях ионов Na и Ca. При интенсивной импульсации выброшенный из везикул пресинаптической мембраны ацетилхолин связывается с рецепторными белками постсинаптической мембраны, подавляет деятельность ионных насосов, деполяризует мембрану и выключает Na+, K+-АТФ-азу. Однако возникающие при этом потенциалы возбуждения и уменьшение отношения Na+ и K+ приводят к высвобождению ацетилхолина и последующему разрушению его холинэстеразой, что может восстанавливать активность Na+, К+-АТФ-азы на более высоком уровне.
Временное повышение синаптической проводимости связывают также с высвобождением Ca2+ под действием нервных импульсов в пресинаптических окончаниях, который вызывает усиленное сокращение везикул и выбрасывание медиатора, или с повышенной чувствительностью белков деполяризованной мембраны к протеолитическим ферментам, делающим ее более проходимой для медиатора. В пользу последнего предположения свидетельствует возрастание активности протеаз в ткани мозга животных при выработке условного рефлекса.
Исходя из результатов электрофизиологических исследований было высказано мнение, что краткосрочная память образуется за счет посттетанической потенциации, поддерживаемой реверберующими потоками импульсов. Посттетанической потенциацией, или посттетаническим облегчением, называют увеличение числа проходимых синапсов после ритмического раздражения пресинаптических волокон. На то, что это явление зависит от изменений в их окончаниях, а не в постсинаптических структурах, указывает отсутствие такого эффекта при антидромной стимуляции (рис. 22). Из рис. 22 видно, что в данном случае облегчение длится около двух минут и что антидромная стимуляция оказывается безрезультатной.
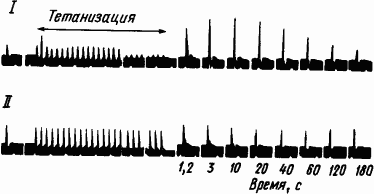
Рис. 22. Посттетаническое временно?е облегчение проведения через звездчатый ганглий кошки (по М. Ларраби, Д. Бронк):
I — ортодромная стимуляция, II — антидромная стимуляция, объяснение см. в тексте
Несмотря на различия предположений о природе краткосрочной памяти, их объединяет рассмотрение механизма памяти как возникновения непродолжительных обратимых физико-химических свойств мембран и динамики медиаторов в синапсах, временно перестраивающих поведение нервных сетей. Эти изменения неизбежно связаны со сложной цепочкой далеко идущих, сравнительно более стойких перестроек метаболизма нервной клетки. Поэтому кратковременная память может оставлять более длительный скрытый след, основе которого в некоторых случаях она способна восстанавливаться после действия амнестического агента. Углубление и структурно-метаболическое закрепление этих следов, по-видимому, является механизмом перехода краткосрочной памяти в долгосрочную.
Долгосрочная память
В отличие от краткосрочной долгосрочная память хранит следы пережитых событий неопределенно долго. Собственно она и является носителем жизненного опыта животного; знаний, приобретенных человеком. О субстрате и способе закрепления долгосрочных памятных следов существуют разные мнения.
Долгосрочная память как возникновение новых межклеточных связей и улучшение проведения по уже существующим связям. Не вызывает сомнений, что в основе всякого обучения лежит процесс проторения путей возбуждения. Наглядной иллюстрацией того, что в результате обучения потоки нервных импульсов легче и быстрее преодолевают синаптическую задержку, может служить модельный опыт с тараканом: после ампутации передних лапок, которыми он чистит усики, таракан обучается пользоваться для этой цели задними лапками. При этом время синаптической задержки моторных импульсов к задним лапкам резко уменьшается (рис. 23).
Исследование динамики активности нейронов виноградной улитки при выработке оборонительного условного рефлекса показало, что повышение эффективности синапсов происходит на входах командных нейронов, реализующих защитные движения.
При выработке условного рефлекса постановки лапы кошки на подставку исследовали реакции ЭЭГ и нейронной активности в моторной коре. Наблюдаемое при этом их единообразие в случае применения различных по характеру и интенсивности условных раздражителей привело к заключению, что сигнал лишь запускает программу ответного двигательного акта. Это происходит, когда наступает повышение эффективности синапсов на входах нейронов — генераторов команд.
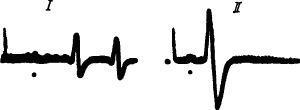
Рис. 23. Укорочение времени прохождения через метаторакальный ганглий таракана нервных импульсов, приводящих в движение его задние лапки, которыми он научился чистить усики после ампутации передних лапок (по Д. Люко, А. Аранда):
I — до ампутации, II —через 12 дней после ампутации, точки под записями — момент прихода импульсов к ганглию, двухфазные потенциалы зарегистрированы от эфферентных волокон данного ганглия
Давно высказанное предположение, что памятный след закрепляется «прорастанием» дополнительных нервных окончаний (Р. Кахал, 1911), позже получило новые подтверждения. Оказалось, что так называемые шипики, несущие синапсы на дендритах корковых клеток, появляются именно в возрасте, когда развивается условно-рефлекторная деятельность. Дополнительные указания на роль шипиков в мехаизмзах памяти дали эксперименты, показавшие интенсивное развитие их синаптического аппарата у животных в результате выработки условных рефлексов. К такому пониманию природы долгосрочной памяти приводит и концепция участия генетического аппарата в пластическом обеспечении функций нервных клеток, согласно которой их интенсивная деятельность при обучении вызывает усиление синтеза белка и процессов роста, в частности, образующих новые синаптические связи. Наконец, зависимость количества синапсов (по показателям электронной микроскопии) от интенсивности функционирования нервных механизмов продемонстрирована в экспериментах с повышением активности синапсов путем предъявления сложных задач или ее снижения путем сенсорной депривации.
По другим предположениям долгосрочная память реализуется облегчением передачи импульсов по существующим синапсам. Это может быть достигнуто стойкими изменениями структуры и свойств синапсов (табл. 3).
Эффективность передачи возбуждения в синапсах в значительной мере зависит от их функционального состояния, определяемого уровнем деятельности. Показано, что усиленное использование синапсов повышает их проходимость для импульсных потоков. При объяснении этого явления учитывается, что ничтожные просветы синаптических щелей соизмеримы с размерами макромолекул мембран, образующих их стенки. Поэтому многократная конвергенция импульсов условного и безусловного раздражителей на синапсах центрального нейрона, вызывая физико-химические изменения его мембран, может, в конечном итоге, сделать синаптическую щель более преодолеваемой.
Физико-химические изменения мембран, возникающие при стойком сдвиге их поляризации, могут служить основой для формирования памятных следов. Такие сдвиги поляризации вопроизведены в экспериментах с созданием зон повышенной возбудимости в коре при помощи постоянного тока и, видимо, имеют место в естественных условиях возникновения доминантных очагов. Опытным путем показано, что создаваемое в доминантном очаге коры состояние возбудимости подпорогового уровня под действием приходящего афферентного возбуждения может достичь порога проявления ответной реакции. Образуется суммационный рефлекс, который при закреплении следов проторения новых путей становится настоящим условным рефлексом. Характерная особенность такого рефлекса состоит в том, что по показателям нейронной активности он протекает в две фазы: торможения и последующей отдачи. Образование таких временных связей происходит преимущественно в нейронах нижних слоев коры.
Одним из механизмов повышения эффективности синапсов могут быть процессы, проявляющиеся в так называемой посттетанической потенциации (Д. Ллойд, 1949). После раздражения афферентных путей синаптическая проводимость мотонейронов спинного мозга остается повышенной в течение 7 мин. Раздражением гиппокампа удавалось продлить такое состояние до нескольких часов. Показано, что гиппокампальная посттетаническая потенциация сопровождается увеличением фокальных потенциалов и укорочением их латентного периода, особенно выраженных, если тетанизации подвергались структуры со свойствами самостимуляции.
Таблица 3. Возможные механизмы устойчивого повышения проводимости в синапсах (по С. Барондес в модификации И.П. Ашмарина)
1. Область изменений в синапсе
2. Состояние синаптической щели
3. Характер изменений
4. Молекулярный механизм
1. Пресинаптическая мембрана
2. Сужение
3. Разрастание нервных окончаний
4. Усиленный синтез белков при запоминании, в дальнейшем их обновление
1. —
2. Без изменения
3. Усиление синтеза и выхода медиатора
4. Дерегрессия части генов медиатора, повышение активности систем трансляции и РНК медиатора
Усиление синтеза медиатора и облегчение выхода из везикул
1. Постсинаптическая мембрана
2. Без изменений
3. Увеличение числа и чувствительности рецепторов
4. Усиленный синтез белков и липидов при запоминании, в дальнейшем их обновление
1. —
2. —
3. Повышение проницаемости и уменьшение скорости расщепления медиатора
4. Измерение соотношений скоростей синтеза компонентов или образование новых компонентов рецептора при запоминании, в дальнейшем поддержание новых отношений. Репрессия генов энзима, расщепляющего медиатор, дерепрессия генов ингибитора этого энзима
1. Локализация не установлена
2. Не известно
3. Повышение проводимости синапса за счет включения неизвестного вещества
4. Усиленный синтез веществ типа антител или других факторов межклеточного «узнавания» (например, полипептиды — коннекторы), при запоминании, в дальнейшем их обновление
Усиленный синтез некоторых антигенов компонента синапса, вызывающих образование соответствующих антител при запоминании, в дальнейшем их обновление
Анализ этих явлений нейронной активности при выработке локальной условной реакции вздрагивания приводит к выводу о том, что для синаптического проторения путей временной связи необходима кроме конвергенции сигнального и безусловного стимулов активация системы, вызывающей модификацию синапсов, что зависит от биологической значимости образования условной реакции. На изолированном нейроне виноградной улитки получены условные изменения свойств его электровозбудимой мембраны путем сочетания стимулов, приложенных к двум ее локусам, в результате чего между ними возникала временная связь.
При изучении возможных причин стойкого повышения синаптической передачи было обращено внимание на медиаторы возбуждения и прежде всего на ацетилхолин. Найдено, что обучение сопровождается повышением активности фермента холинэстеразы, разрушающей ацетилхолин, а вещества, подавляющие действие холинэстеразы, вызывают нарушения памяти. В опытах с обучением крыс доставать пищу из узкой трубки определенной передней конечностью активность холинэстеразы возрастала только в контралатеральном полушарии, что подтверждает ее значение в формировании памятных следов.
Сопоставление данных о взаимосвязанной динамике ацетилхолина и холинэстеразы привело к заключению, что повышенная проводимость может возникать и устойчиво поддерживаться некоторым равновесным отношением их синтеза. Избыточная активность холинэстеразы без возрастания количества ацетилхолина не позволяет ему достичь концентрации, необходимой для деполяризации постсинаптической мембраны синапса, а увеличение количества ацетилхолина при малой активности холинэстеразы приводит к глубокой неколеблющейся деполяризации — в обоих случаях проведение импульсов через синапс оказывается блокированным. Существует предположение, что ацетилхолин может транспортироваться через синаптическую щель по транссинаптическим тяжам, которые образуются из соединения белковых мицелл, формирующихся на поверхности пре- и постсинаптической мембран.
Кроме холинэргических имеются и иные синапсы, где медиаторами служат другие вещества. С учетом всех этих сведений была составлена обобщенная схема стойкого облегчения медиаторных процессов в синапсе. На рис. 24, А показано исходное состояние синапсов. Если возбуждение пресинаптического нейрона (рис. 24, Б) вызывает усиленное выделение медиатора, а возбуждение постсинаптического — разрушающего медиатор фермента (рис. 24, В), то при сочетаниях условного и безусловного раздражителей происходит конвергенция импульсных потоков на синапсах (рис. 24, Г), которая обусловливает оптимальный баланс активности медиатора и разрушающего его фермента на стабильном высоком уровне, а также активацию сети белковых тяжей, уменьшающих просвет синаптической щели (рис. 24, Д). Показано, что повышение эффективности синапсов сопровождается конформационными изменениями мембранных белков, специфичными для синаптических мембран.
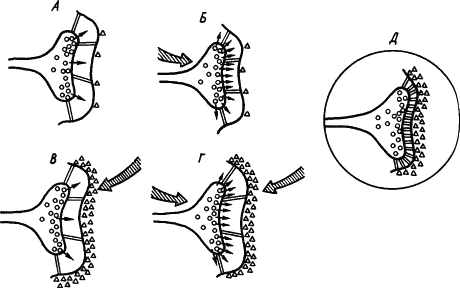
Рис. 24. Развитие процессов стойкого облегчения проводимости сигнала (объяснение см. в тексте) (по В.В. Дергачеву):
кружками обозначены везикулы медиатора, треугольниками — фермент, расцепляющий медиатор
Долгосрочная память как стойкое изменение химизма нервных клеток. Проведение сигналов от клетки к клетке, вызывающее активацию нейронных популяций, происходит в синапсах, однако функциональное состояние синапсов и свойства их деятельности в значительной мере определяются процессами, происходящими в соме клетки, в ее нейроплазме и ядре. Общеизвестна роль сомы в трофическом обеспечении передачи импульсов по путям их проведения и зависимости функционирования нейрона от его метаболизма. Это послужило основанием для поисков долгосрочного памятного следа в стойких изменениях химизма нейроплазмы.
Так как основным субстратом функциональных структур нейрона являются белковые соединения, а их различные модификации задаются матрицами РНК, на которых они синтезируются, то была выдвинута гипотеза о том, что информация, входящая в долгосрочную память, «записывается» структурой полинуклеотидной цепи молекулы (X. Хиден, 1959). По этой гипотезе разные условные потоки импульсов создают различное распределение ионных концентраций вдоль молекулы РНК и вызывают специфические для каждого сигнала перемещения нуклеотидов в их цепи. В результате каждый сигнал получает определенный отпечаток в структуре молекулы РНК. Поскольку структура молекулы РНК является матрицей для синтеза белковых соединений, специфические белки и медиаторы могут синтезироваться лишь по тому сигналу, который сформировал эту матрицу своим импульсным потоком. Все возможные перестановки и комбинации нуклеотидных элементов позволяют зафиксировать в молекулах РНК громадное количество информации. По приблизительному подсчету их информационная емкость достигает 1015–1020 бит, что перекрывает объем человеческой памяти.
В качестве экспериментального подтверждения гипотезы о переопределении нуклеотидов в молекуле РНК как механизме формирования долгосрочной памяти могут служить результаты опытов с крысами, которые обучались добираться к пище, балансируя на проволочном мостике (табл. 4).
Таблица 4. Процентное содержание нуклеотидов в РНК ядра Дейтерса при выработке двигательного навыка у крыс (по X. Хидену, Е. Эльхази)
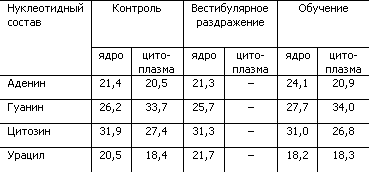
Как показывает табл. 4, у крыс, обучившихся сложным движениям, в нервных клетках ядра Дейтерса, ответственного за выполнение нового двигательного навыка, произошло изменение нуклеотидного состава РНК клеточного ядра. Увеличилась доля аденина, уменьшилась доля урацила. Простое вестибулярное раздражение не изменяло нуклеотидный состав. Исследование глиальных клеток ядра Дейтерса выявило сходные изменения РНК, что позволило сделать предположение об участии глии в кодировании поступающих сигналов путем изменения структуры нуклеотидных цепей РНК.
Однако гипотеза о нуклеотидном коде памятного следа встретилась со многими затруднениями, из которых главное — недолговечность молекул РНК. Выполняя информационные и транспортные функции, они могут сохранять свою структуру лишь в течение короткого времени, не соизмеримого с длительностью долгосрочной памяти. Эти и другие возражения ставят под сомнение РНК как носителя долгосрочного памятного следа, с чем, в сущности, согласился сам автор нуклеотидной гипотезы (X. Хиден, 1969).
В связи с этим внимание исследователей обратилось к более долговечным химическим компонентам структурно-функциональной организации нервной клетки. Памятный след стали искать в генетическом аппарате, который управляет деятельностью нервной клетки и определяет свойства на протяжении всей ее жизни. Наиболее стабильной структурой генетического аппарата являются молекулы ДНК, в которых кодируется наследственная информация. Логично было думать, что и информация, приобретаемая в течение жизни фиксируется этими структурами, специально приспособленными к хранению информационных программ. Поэтому были предприняты исследования ДНК как носителя долговременной памяти.
Анализ свойств ДНК и влияния ингибиторов ее синтеза на память привел к предположению, что формирование памятного следа связано с активацией определенных генов при деспирализации соответствующих участков цепи ДНК. Это происходит под влиянием ионных сдвигов, отражающих возбуждение нервной клетки при поступлении в нее сигналов.
На основании того, что снятие гистоновой блокады дерепрессирует гены, обусловливающие модификацию свойств клонов клеток была предложена нейроселективная теория долговременной памяти (В.В. Дергачев, 1967). Согласно этой теории при запоминании происходит селективная дерепрессия генов, реализующая определенную комбинацию из закодированных в геноме клонов нейроглиальных комплексов. В этих комплексах более лабильная функциональная ДНК программирует такое направление синтеза РНК и белков, которое определяет специфические свойства избирательной возбудимости нервных клеток. О значении ДНК в формировании долгосрочной памяти свидетельствуют опыты, показывающие нарушения памяти при действии ингибиторов синтеза рибонуклеиновых кислот даже в то время, когда синтез белков продолжался.
Электрофизиологические исследования механизмов привыкания нервных клеток к действию раздражителей привели к представлению, что памятный след отражает не столько временные, сколько пространственные характеристики сигналов. Он представляет собой не «паттерн импульсов», а «паттерн синапсов», кодирующих своим расположением и состоянием значение сигнала (Е.Н. Соколов, 1969). При этом медиаторы через рецепторы синаптических мембран вызывают активацию одних и инактивацию других оперонов молекулы ДНК. В результате формируется система обеспечения синтеза специфических белков, реализующих функцию «обученной нервной клетки (рис. 25).
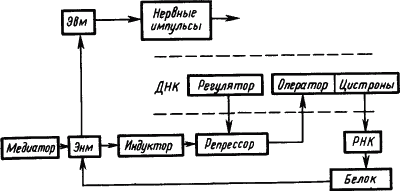
Рис. 25. Участие функциональных структур генетического аппарата молекулы ДНК в долговременной памяти (по Е.Н. Соколову):
пунктиром обозначен участок молекулы ДНК; Эвм — электровозбудимая мембрана, Энм — электроневозбудимая мембрана
За последнее время получены новые доказательства участия ДНК в механизмах долговременной памяти. Показано, что при выработке условных рефлексов происходят своеобразные изменения структуры молекулы ДНК в нервных клетках мозга крыс. Эти изменения состояли в увеличении степени метилирования ДНК, повышающего активность определенных генов. Они были четко выражены в коре больших полушарий и гиппокампе и оказались незначительными в мозжечке. При такой «мягкой» обратимой модификации ДНК ее нуклеотидный состав оставался неизменным (табл. 5).
Таблица 5. Содержание нуклеотидов и метиловых компонентов в ДНК из разных отделов мозга у обученных и контрольных крыс (по Н.А. Тушмаловой)

Как уже было упомянуто, основные структуры нервной клетки, реализующие ее функции, состоят главным образом из белковых соединений. Поэтому многие исследователи ведут изучение белков как возможного субстрата долговременной памяти. Усиленный синтез белка при обучении дает основания для суждений о его роли в явлениях памяти. Чтобы выяснить, какие именно белки усиленно синтезируются для обеспечения процессов фиксации памятного следа, предпринималось изучение их раздельных фракций.
Исследование белков мозга голубей выявило, что при их обучении происходит возрастание фракции мембранных гликопротеидов, обозначенных 10B и 11A. Эти белки благодаря своим углеводным компонентам могут регулировать активность возбудимых мембран и таким образом участвовать в процессах памяти. При обучении крыс применять непредпочитаемую лапу для добывания пищи последующее дробное фракционирование мозговых белков показало особое положение встречающихся только в нервной ткани белков S-100 и 14-3-2. Конформационные изменения S-100 при связывании с ионами Ca влияют на проницаемость мембран, в частности синаптических, определяя их функциональное состояние.
Значение белков в механизмах формирования долгосрочной памяти продемонстрировано в опытах, где угнетение синтеза белка нарушало переход краткосрочной памяти в долгосрочную. Так, введение ингибитора белкового синтеза пуромицина золотым рыбкам сразу после обучения стирало его следы, но введение пуромицина через 1 ч не вызывало такого действия. Эти опыты были повторены и на других животных, показывая, что синтез белка необходим для консолидации памяти, а когда памятный след закреплен, белок уже не столь нужен.
Какие же именно белки разрушаются под действием ингибитора, препятствующего формированию долговременной памяти? Для ответа на этот вопрос исследовали разные фракции мембранных белков синаптического аппарата после нарушения долговременной памяти пуромицином. Показано, что этот ингибитор наиболее резко тормозит синтез кислых белков, к которым относится специфический для тканей мозга белок S-100. Оказалось также, что белки наиболее чувствительной к ингибитору фракции по своему аминокислотному составу принадлежат к группе внутренних белков мембраны, которые связаны с ее функциями как холинорецепторы. Это соответствует мнению, что носителями долгосрочной памяти являются холинэргические синапсы. Однако имеются указания на участие в механизмах долгосрочной памяти и адренэргической системы. Отмечено, что положительное подкрепление условных рефлексов, например пищевых, связано с функционированием преимущественно серотонинэргических, а отрицательное при оборонительных рефлексах — норадренэргических структур мозга. О возможных нейрохимических компонентах подкрепления существуют разные мнения: сюда относят и норадренэргическую систему, и дофаминэргическую. За последнее время накоплено большое количество фактов, показывающих, как участвуют нейрохимические механизмы в процессах обучения и памяти.
Другой подход к выяснению роли белков в долговременной памяти состоял в прослеживании включения в них меченых аминокислот и изменений этих белков при обучении. Так, внедрение меченного тритием лейцина в белки мозга крыс после обучения возрастало и сопровождалось увеличением числа рибосом, связанных с мембранами. При переучивании крыс способу добывания пищи на стадии бурной двигательной активности происходило даже снижение включения меченого лейцина в белки мозга. По мере усвоения нового навыка включение меченого лейцина в белки, особенно специфические, возрастает. После выработки нового навыка п<...пропуск части слова...>ния у золотых рыбок отмечен синтез трех новых фракций белков, обнаруженных с помощью меченого валина. У крыс, обученных дифференцированию зрительных сигналов для решения лабиринтной задачи, наблюдали ускоренное включение меченого тирозина в мембранные белки. Выработка условных рефлексов у голубей сопровождалась усилением синтеза гликопротеинов по показателям включения меченого валина.
В последние годы все более широко применяют методы иммунохимической идентификации специфичных для мозга белков, в частности S-100, 14-3-2; гликопротеинов для определения их роли в фазировании свойств возбудимых мембран и синаптических процессов. Таким образом, накапливаются веские доказательства непосредственного участия белков, особенно кислых мембранных и гликопротеинов в переходе краткосрочной памяти в долгосрочную.
Сравнительно недавно было выяснено, что к возможным носителям долгосрочной памяти относится ряд пептидов — веществ, как и белки, состоящих из аминокислот, но более простых по своему строению. Такие пептиды ускоряют формирование условных рефлексов. Их действие на другие организмы воспроизводит выработанный навык, что рассматривается как явление «переноса памяти» (см. ниже).
К этим пептидам относятся главным образом такие гормоны гипофиза, как адренокортикотропин, меланоцитостимулирующий гормон, вазопрессин, окситоцин и некоторые рилизинг-гормоны; а также пептиды, специфически связанные с обучением какому-либо одному определенному навыку. Связь пептидов с памятью, по-видимому, определяется их участием в синаптической передаче. Предполагают даже существование особых «пептидэргических синусов», которые могут изменять свои свойства проведения.
Кроме того, считают, что факторы пептидной природы могут участвовать в образовании временных связей как химический механизм воздействия подкрепляющих систем на генетический аппарат нервных клеток.
Одно из замечательных свойств памяти состоит в специфической избирательности и локальности образуемых временных связей. В хорошо выработанном условном рефлексе каждый сигнал вызывает только свою строго определенную реакцию. В этом, а также в длительности хранения, свойствах угасания и восстановления, в чувствительности к ядам, тормозящим синтез белков и РНК, «память» нервной системы очень напоминает «память» системы иммунитета. Предположение об участии иммунных реакций в формировании долгосрочной памяти получило новые подтверждения и привело к разработке гипотезы об иммунохимическом механизме формирования долговременной памяти (И.П. Ашмарин, 1973).
Согласно этой гипотезе в результате метаболизма синаптических мембран на их поверхности образуются вещества, которые, накапливаясь в избытке, при реверберации импульсов в стадии краткосрочной памяти играют роль антигена для антител, вырабатываемых в глиальных клетках. Связывание антитела с антигеном происходит при участии активаторов образования медиаторов или ингибитора ферментов, расщепляющих эти активаторы (рис. 26). В пользу этой гипотезы свидетельствуют эксперименты, показавшие, что антитела к синаптосомам повышали чувствительность последних к медиаторам и что стимуляторы иммуногенеза улучшали долгосрочную память.
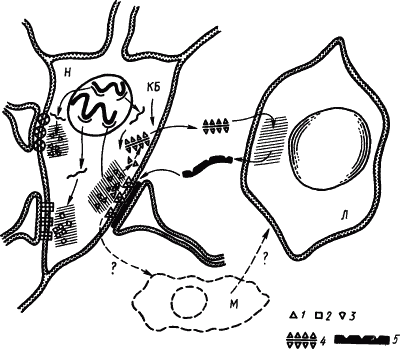
Рис. 26. Иммунологические механизмы, повышающие проводимость определенных синапсов (по И.П. Ашмарину):
КБ — катионные белки, открывающие путь комплексам фрагментов антигена синапса с РНК через оболочку клетки; Л — клетки глии, функционально подобные лимфоцитам, М — клетки глии, функционально подобные макрофагам; 1, 2, 3 — фрагменты синаптических антигенов (зоны их синтеза заштрихованы), 4 — комплекс фрагмента антигена с РНК, 5 — антитела для синапсов, тонкими линиями изображена мРНК, толстыми — ДНК хромосом
Участие в иммунохимических механизмах памяти специфичного для мозга белка S-100 показано в опытах эпидурального введения крысам сыворотки с антителами к этому белку, в результате чего затруднялась выработка навыка поведения в лабиринте. Чтобы выделить в «чистом виде» эффект действия антител на белки, специфические для процессов образования временных связей, изучали навык поведения крыс. При этом вводили антитела, которые вырабатывались на гомогенаты мозга необученных и обученных крыс, а также крыс, получавших препараты, облегчающие или затрудняющие обучение.
Эффект действия антител резко проявлялся уже в 1-й день обучения в виде угнетения условных реакций животных, получавших антитела к веществам-участникам формирования долговременной памяти под влиянием препаратов, облегчающих обучение. Видимо, такой эффект объясняется связыванием и дезактивацией этих веществ.
Больше половины мозга занимают глиальные клетки, которых примерно в 10 раз больше, чем нервных; каждый нейрон со всех сторон окружен глией. Естественным было предположение, что глия участвует в осуществлении функций мозга, в том числе высшей нервной деятельности. Замедленность реакций клеток глии и сосредоточение в них специфичных для мозга белков S-100, некоторых гликопротеинов и биологически активных соединений создают условия для реализации постепенно стабилизирующихся процессов, к каким относится память. Поэтому появились основания предполагать, что клетки глии во взаимодействии с нейронами принимают деятельное участие в формировании долгосрочной памяти.
Для подтверждения этого предположения проводили эксперименты, показавшие, что инъекции экстрактов глии крысам ускоряли выработку условных рефлексов. В пользу такого предположения свидетельствует и прогрессивное увеличение глиальной ткани относительно нервной в ряду от низших животных к высшим. Медленная электрическая активность глии может быть источником колебаний постоянного потенциала мозга и электротонических потенциалов, создающих очаги повышенной возбудимости, которые считают основой образования временных связей.
О возможном механизме участия глии в фиксации памятного следа существуют различные мнения. Так, предполагали, что выходящие при возбуждении нейрона ионы K захватываются клетками глии и передаются следующему нейрону, вызывая стойкое изменение уровня его поляризации, а следовательно, облегчение его возбуждения. Другое объяснение состоит в том, что выработка условного рефлекса вызывает у прилегающих к нейрону глиальных клеток усиленное образование миелина, который окутывает его тончайшие терминальные волокна и уменьшает фактор риска проведения по ним импульсов, в результате чего «потенциальные» синапсы становятся действующими (А.И. Ройтбак, 1968). Схема происходящих при этом процессов представлена на рис. 27. Эта гипотеза получила экспериментальное подтверждение в стимуляции миелинообразования глиальными клетками с помощью ионов K, которые выходят из возбужденного нейрона в межклеточную среду.
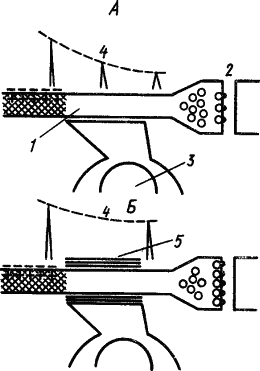
Рис. 27. Гипотеза о роли глии в образовании временной связи условного рефлекса. А — синапс не проводит, так как потенциал возбуждения в немиелинизированной пресинаптической терминали ослабляется и медиатор выделяется в количестве, недостаточном для срабатывания синапса; Б — глиальная клетка, возбуждаясь при безусловном подкреплении рефлекса, замыкает пресинаптический участок терминали в миелиновую оболочку и потенциал возбуждения сохраняет величину, необходимую для срабатывания синапса (по А.И. Ройтбаку):
1 — аксон, 2 — синапс, 3 — глиальная клетка, 4 — изменение потенциала действия, 5 — миелиновая оболочка
Для того чтобы определить, какова в долговременной памяти роль глии и нейронов, испытывали действие на условные рефлексы крыс антител, полученных при иммунизации гомогенатом или глии, или нейронов. Оказалось, что антинейрональная сыворотка ухудшала главным образом краткосрочную память, а антиглиальная — долгосрочную. Эти результаты рассматривают как указание на то, что краткосрочная память обусловлена реверберацией импульсов по цепям нейронов, а долгосрочная — реализуется пластическими процессами с участием глии.
Уже давно высказали мнение, что нервные клетки выполняют свои функции в виде объединений. Таковы динамические констелляции (А.А. Ухтомский, 1945) и образующиеся при обучении функциональные ансамбли (Д. Хебб, 1949). Экспериментальные исследования показали реальность существования таких объединений, названных элементарными нейронными ансамблями (А.Б. Коган, 1964, 1979). Нейронные ансамбли обладают свойствами, отличными от свойств отдельных нейронов.
Качественное своеобразие системной деятельности нейронов обосновывается морфологическими особенностями их связей (Я. Сентаготаи, 1968), эффектами объединения; существует представление о том, что при обучении временные связи формируются путем объединения нейронов различных структур мозга в комплексы, называемые микросистемами (М.М. Хананашвили, 1972). Наименьшие микрообъединения нейронов коры, строящие свою деятельность на основе предыдущего опыта, обозначают как нейроценозы. Локальная аппликация ацетилхолина в качестве подкрепления выявляла группы нейронов, проявляющие синхронные пластические перестройки. Эти нейроны определяли как микропулы.
Существует концепция, по которой каждый акт высшей нервной деятельности связан с активацией определенной совокупности нейронов, образующей сложную мозаику. Такой подход к проблеме памяти лишен затруднений, возникающих перед гипотезами ее «молекулярного кодирования», и подкрепляет концепцию «распределенной памяти», согласно которой механизмы фиксации памятного следа не сводятся к изменениям свойств отдельных клеток, а состоят в избирательном формировании определенных межнейронных отношений в их популяциях. Об этом свидетельствуют и результаты опытов с так называемым обучением по обратным связям, когда подкреплялась импульсация одного из трех одновременно регистрируемых нейронов слуховой коры кошки. Корреляционный анализ показал, что нейрон, импульсация которого служила сигналом, устанавливал определенную систему связи с остальными нейронами и такие межнейронные отношения могли сохраняться до 20 мин. Представление о совместной деятельности нейронных объединений легло в основу теории статистической конфигурации (Е. Джон, 1973), согласно которой носителем памятного следа является статистически формируемый набор активных нейронов, образующих фигуру определенных очертаний.
Ha основании большого экспериментального материала о взаимосвязи ритмических свойств нейронов, образующих цепочки возбуждения разной лабильности и определяющих функциональное состояние центральных нервных структур, было высказано мнение, что новая временная связь запечатлевается в нервной системе в виде специфического пространственно-временного распределения активности нейронов, перестроивших свою лабильность (М.Н. Ливанов, 1975). В пользу этого предположения свидетельствуют результаты исследования конфигурации мозаики нейронной активности, показавшие, что условный рефлекс проявляется лишь тогда, когда рисунок мозаики, вызванной сигналом, соответствует таковому памятных следов и подходит к нему, как «ключ к замку». Возникновение разных рисунков нейронной активности с участием одних и тех же нейронов означает перераспределение свойств проводимости их синапсов, которое может осуществляться рассмотренными выше механизмами.
Такое представление близко к взглядам на нейронную структуру условного рефлекса, как на массовое объединение нейронов, в котором воспроизводится вся ситуация в целом (А. Аттли, Т. Блумфилд, Е. Джон, 1973). Обобщение большого материала многолетних нейрохимических исследований памяти также привело к заключению о ее распределенном характере в ансамблях нейронов. При этом роль биохимических процессов, создающих сложные «рисунки» распределения химизма нервных клеток, заключается в организации «узнавания» клетками друг друга и «сборки» их в функциональные ансамбли. Существует предположение о химической природе сигнала, управляющего проходимостью синапсов нейронов. Это выражается перестройкой классов доминирующих межимпульсных интервалов, в активности нейронов.
Получены также факты, указывающие на то, что механизм образования условного рефлекса не ограничивается перестройками центральных процессов, а включает в себя и периферические гормональные компоненты. Особое значение при этом имеет система гипофиз — надпочечники и образование энкефалиноподобных пептидов. Эксперименты с действием мет- и лей-энкефалина на условные рефлексы избегания у крыс привели к предположению об их возможной роли в обучении и памяти.
«Перенесение» памяти
Особое место в исследованиях занимают явления, получившие название перенесение памяти. Начало им положило сообщение об опытах на плоских червях (планариях), которых кормили планариями с предварительно выработанным двигательным защитным рефлексом. При этом у кормленных таким образом червей наблюдали ускоренную выработку условного рефлекса.
Результаты экспериментов по изучению условных реакций планарий, получавших экстракт РНК от «обученных» и «необученных» особей, дали основание предполагать, что «переносчиком» памяти может служить именно РНК.
Опыты проводились и на позвоночных животных, которым вводили экстракт мозга «обученных» животных. Так, экстракт мозга рыбок, у которых был выработан условный рефлекс избегания ударов тока по сигналам синего или зеленого цвета, при введении его необученным рыбкам вызывал у них отрицательную реакцию на соответствующий цвет. Многочисленные опыты с «переносом» памяти были поставлены на крысах, причем были получены как положительные, так и отрицательные результаты (например, исследование влияния такого экстракта мозга на выработку целевых условных рефлексов).
Поиски активного начала мозговых экстрактов привели к выделению из них пептидов, обнаруживших высокую избирательность своего действия (Г. Унгар, 1972). Так, из экстракта мозга крыс, обученных избегать темноту (его введение вызывало у необученных также избегание темноты), был выделен пептид, состоящий из 15 аминокислот. Он получил название скотофобин (вызывающий боязнь темноты). Скотофобин был синтезирован и проявлял такое же действие, как естественный. У животных, оказавшихся в темноте, он вызывал реакцию стресса.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 9 Что такое примат? Развитие со времен динозавров. Кометы и прочие эволюционные механизмы.
Глава 9 Что такое примат? Развитие со времен динозавров. Кометы и прочие эволюционные механизмы. Как я уже говорил, палеонтологический «пробел» измеряется четырьмя миллионами лет – целых сорок метров на нашей мерной ленте, где один миллиметр соответствует ста
§ 15. Механизмы памяти
§ 15. Механизмы памяти 1. Память — это функция нервных клеток. При синдроме Корсакова, рассеянном склерозе, ишемической болезни мозга, когда дегенерируют нейроны, память исчезает.2. Для способности запоминать надо иметь некоторое количество нейронов, по-видимому, немного
1.4. Возрастные анатомо-физиологические особенности
1.4. Возрастные анатомо-физиологические особенности Для каждого возрастного периода характерны количественно определенные морфологические и физиологические показатели. Измерение морфологических и физиологических показателей, характеризующих возрастные,
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции Пер. А. НестеровойВ этой главе мы продолжим обсуждение эволюционной биологии в период до появления геномики. Многие из обсуждаемых направлений развития не являлись
Глава 6 ПСИХОФИЗИОЛОГИЯ ПАМЯТИ
Глава 6 ПСИХОФИЗИОЛОГИЯ ПАМЯТИ Научение может быть рассмотрено как последовательность сложных процессов, вовлекаемых в приобретение, хранение и воспроизведение информации (см. гл. 15). В результате научения происходит модификация поведения, а память проявляется как
5. МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ПАМЯТИ
5. МОЛЕКУЛЯРНЫЕ МЕХАНИЗМЫ ПАМЯТИ В нейронауках исследование механизмов научения и памяти ведётся преимущественно в контексте пластичности (см. гл. 15). Именно поэтому многие исследования имели своей целью идентификацию пластических изменений активности и морфологии
Способы вызывания гипноза: физиологические и психические приемы
Способы вызывания гипноза: физиологические и психические приемы Обращаясь к вопросу о вызывании гипноза, необходимо иметь в виду, что для этого с пользою могут служить различные способы, из которых одни могут считаться физиологическими, так как они действуют
ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ СНА
ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ СНА Природа сна постоянно интересует врачей, ученых разных специальностей-биологов, психологов, философов, да и простых людей. Величайшие мыслители уже давно обсуждали эту проблему. Великий врач древности Гиппократ полагал, что сон возникает в
Глава 4 ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ РЕГУЛЯЦИИ ВЕГЕТАТИВНЫХ ФУНКЦИЙ И ИНСТИНКТИВНОГО ПОВЕДЕНИЯ
Глава 4 ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ РЕГУЛЯЦИИ ВЕГЕТАТИВНЫХ ФУНКЦИЙ И ИНСТИНКТИВНОГО ПОВЕДЕНИЯ К вегетативным относят те функции, которые обеспечивают обмен веществ в нашем организме (пищеварение, кровообращение, дыхание, выделение и др.). К ним относят также обеспечение
3.5. Механизмы памяти
3.5. Механизмы памяти Поведение высших животных было бы невозможно без использования прошлого опыта, т.е. без сохранения информации о нем в нервной системе. Сенсорная память проявляется в удержании в нейроне следа от раздражения рецептора на очень короткое время — до 0,5 с.
Глава 2. ЭКЗОТРОФИЯ. МЕХАНИЗМЫ ПИЩЕВАРЕНИЯ
Глава 2. ЭКЗОТРОФИЯ. МЕХАНИЗМЫ ПИЩЕВАРЕНИЯ Метаболический цикл начинается с поступления веществ, необходимых для выполнения живыми системами их основных функций. Все живые организмы нуждаются в пище, т.е. источниках энергии, строительных и пластических материалах, солях
Первый отдел Физиологические цвета
Первый отдел Физиологические цвета 1 Эти цвета, которые мы справедливо ставим на первое место, потому что они частью полностью, частью преимущественно принадлежат субъекту, глазу его; эти цвета, которые образуют основу всего учения и раскрывают нам хроматическую
Глава 3 Механизмы зарождения жизни на Земле
Глава 3 Механизмы зарождения жизни на Земле 3.1. Аминокислоты Сформировавшиеся физико-химические условия на первобытной планете можно отождествить с установкой С. Миллера, в которой он синтезировал аминокислоты из газов, существовавших в тот период. Единственная разница
Глава 4 МЕХАНИЗМЫ РАБОТЫ ЯЗЫКА Грамматика за работой
Глава 4 МЕХАНИЗМЫ РАБОТЫ ЯЗЫКА Грамматика за работой Журналисты говорят, что это не новость, когда собака кусает человека; вот когда человек кусает собаку — это что-то новенькое. И именно в этом заключена суть языкового инстинкта: язык сообщает о чем-то новом. Цепочки
1. ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ РАЗГРУЗОЧНО-ДИЕТИЧЕСКОЙ ТЕРАПИИ
1. ФИЗИОЛОГИЧЕСКИЕ ОСНОВЫ РАЗГРУЗОЧНО-ДИЕТИЧЕСКОЙ ТЕРАПИИ Под голоданием следует понимать состояние организма при полном или недостаточном поступлении пищевых веществ, а также выраженных нарушениях их состава или усвоения.Голодание может быть полное («влажное»),