Глава 2 ОБРАЗОВАНИЕ УСЛОВНЫХ РЕФЛЕКСОВ
Глава 2
ОБРАЗОВАНИЕ УСЛОВНЫХ РЕФЛЕКСОВ
Основной элементарный акт высшей нервной деятельности — образование условного рефлекса. Здесь эти свойства будут рассмотрены, как и все общие законы физиологии высшей нервной деятельности, на примере условных слюноотделительных рефлексов собаки.
Условный рефлекс занимает высокое место в эволюции временных связей, являющихся универсальным приспособительным явлением в животном мире. Наиболее примитивный механизм индивидуального приспособления к меняющимся условиям жизни, видимо, представляют внутриклеточные вре?менные связи простейших. У колониальных форм развиваются зачатки межклеточных вре?менных связей. Возникновение примитивной нервной системы сетчатого строения дает начало вре?менным связям диффузной нервной системы, обнаруживающимся у кишечнополостных. Наконец, централизация нервной системы в узлы беспозвоночных животных и мозг позвоночных приводит к бурному прогрессу вре?менных связей центральной нервной системы и возникновению условных рефлексов. Столь разные типы вре?менных связей, очевидно, осуществляются физиологическими механизмами различной природы.
Существует бесчисленное множество условных рефлексов. При соблюдении соответствующих правил любой воспринимаемый раздражитель можно сделать стимулом, запускающим условный рефлекс (сигналом), а любую деятельность организма — его основой (подкреплением). По роду сигналов и подкреплений, а также по отношениям между ними созданы разные классификации условных рефлексов. Что касается изучения физиологического механизма временных связей, то здесь исследователям предстоит большая работа.
Общие признаки и виды условных рефлексов
На примере систематического изучения слюноотделения у собак наметились общие признаки условного рефлекса, а также частные признаки разных категорий условных рефлексов. Определилась классификация условных рефлексов по следующим частным признакам: 1) обстоятельства образования, 2) род сигнала, 3) состав сигнала, 4) род подкрепления, 5) отношение во времени условного раздражителя и подкрепления.
Общие признаки условных рефлексов. Какие признаки являются общими и обязательными для всех условных рефлексов? Условный рефлекс а) является индивидуальным высшим приспособлением к меняющимся условиям жизни; б) осуществляется высшими отделами центральной нервной системы; в) приобретается путем вре?менных нервных связей и утрачивается, если вызвавшие его условия среды изменились; г) представляет собой предупредительную сигнальную реакцию.
Итак, условный рефлекс — это приспособительная деятельность, осуществляемая высшими отделами центральной нервной системы путем образования вре?менных связей между сигнальным раздражением и сигнализируемой реакцией.
Натуральные и искусственные условные рефлексы. В зависимости от характера сигнального раздражителя условные рефлексы делят на натуральные и искусственные.
Натуральными называют условные рефлексы, которые образуются в ответ на воздействие агентов, являющихся естественными признаками сигнализируемого безусловного раздражения.
Примером натурального условного пищевого рефлекса может служить выделение слюны у собаки на запах мяса. Этот рефлекс с неизбежностью образуется естественным путем в течение жизни собаки.
Искусственными называют условные рефлексы, которые образуются в ответ на воздействие агентов, не являющихся естественными признаками сигнализируемого безусловного раздражения. Примером искусственного условного рефлекса может служить выделение слюны у собаки на звук метронома. В жизни этот звук не имеет никакого отношения к еде. Экспериментатор искусственно сделал его сигналом приема пищи.
Натуральные условные рефлексы природа вырабатывает из поколения в поколение у всех животных соответственно их образу жизни. В результате натуральные условные рефлексы легче образуются, скорее укрепляются и оказываются более прочными, чем искусственные. Щенок, никогда не пробовавший мяса, безразличен к его виду. Однако достаточно ему один-два раза съесть мясо, и натуральный условный рефлекс уже закреплен. При виде мяса у щенка начинается слюноотделение. А для того чтобы выработать искусственный условный рефлекс слюноотделения на вид вспыхивающей лампочки, нужны десятки сочетаний. Отсюда становится понятным значение «биологической адекватности» агентов, из которых делаются раздражители условных рефлексов.
Избирательная чувствительность к экологически адекватным сигналам проявляется в реакциях нервных клеток мозга.
Экстероцептивные, интероцептивные и проприоцептивные условные рефлексы. Условные рефлексы на внешние раздражители называют экстероцептивными, на раздражители от внутренних органов —интероцептивными, на раздражители скелетно-мышечной системы — проприоцептивными.
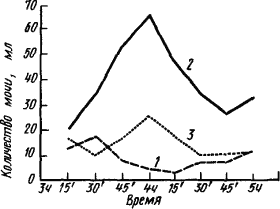
Рис. 1. Интероцептивный условный рефлекс мочеобразования при «мнимом вливании» физиологического раствора (по К. Быкову):
1 — исходная кривая мочеобразования, 2 — мочеобразование в результате вливания в желудок 200 мл физиологического раствора, 3 — мочеобразование в результате «мнимого вливания» после 25 истинных
Экстероцептивные рефлексы делят на рефлексы, вызываемые дистантными (действующими на расстоянии) и контактными (действующими при непосредственном соприкосновении) раздражителями. Далее они разбиваются на группы по основным видам сенсорного восприятия: зрительного, слухового и т.д.
Интероцептивные условные рефлексы (рис. 1) можно также группировать по органам и системам, являющимся источниками сигнализации: желудочные, кишечные, сердечные, сосудистые, легочные, почечные, маточные и т.д. Особое положение занимает так называемый рефлекс на время. Он проявляется в различных жизненных отправлениях организма, например в суточной периодичности обменных функций, в выделении желудочного сока при наступлении времени обеда, в способности просыпаться в назначенный час. По-видимому, организм «отсчитывает время» главным образом по интероцептивным сигналам. Субъективное переживание интероцептивных рефлексов не имеет образной предметности экстероцептивных. Оно дает лишь расплывчатые «темные чувства» (термин И.М. Сеченова), из которых складывается общее самочувствие, отражающееся на настроении и работоспособности.
Проприоцептивные условные рефлексы лежат в основе всех двигательных навыков. Они начинают вырабатываться с первых взмахов крыльев птенца, с первых шагов ребенка. С ними связано овладение всеми видами локомоций. От них зависит слаженность и точность движения. Совершенно новое использование получают проприоцептивные рефлексы руки и голосового аппарата у человека в связи с трудом и речью. Субъективное «переживание» проприоцептивных рефлексов состоит главным образом в «мышечном чувстве» положения тела в пространстве и его членов друг относительно друга. Вместе с тем, например, сигналы от аккомодационных и глазодвигательных мышц имеют зрительный характер восприятия: дают информацию об удаленности рассматриваемого объекта и его перемещениях; сигналы от мускулатуры кисти и пальцев дают возможность оценить форму предметов. С помощью проприоцептивной сигнализации человек своими движениями воспроизводит события, происходящие вокруг него (рис. 2).
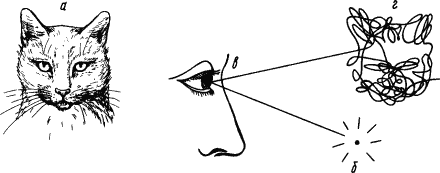
Рис. 2. Изучение проприоцептивных компонентов зрительного представления человека:
а — изображение, ранее показанное исследуемому, б — источник света, в — отражение светового пучка от зеркальца, укрепленного на глазном яблоке, г — траектория движения глаза при воспоминании об изображении
Особую категорию условных рефлексов составляют модельные опыты с электрическим раздражением мозга в качестве подкрепления или сигнала; с использованием в качестве подкрепления ионизирующей радиации; созданием доминанты; выработкой временных связей между пунктами нейронально-изолированной коры; исследованием суммационного рефлекса, а также образованием условных реакций нервной клетки на сигнал, подкрепляемый локальной электрофоретической аппликацией медиаторов.
Условные рефлексы на простые и комплексные раздражители. Как было показано, условный рефлекс можно выработать на какой-либо один из перечисленных экстеро-, интеро- или проприоцептивных раздражителей, например на включение света или на простой звук. Но в жизни это бывает редко. Чаще сигналом становится комплекс из нескольких раздражителей, например запах, тепло, мягкая шерсть матери-кошки становятся раздражителем условного сосательного рефлекса для котенка. Соответственно разделяют условные рефлексы на простые и сложные, или комплексные, раздражители.
Условные рефлексы на простые раздражители не требуют пояснений. Условные рефлексы на сложные раздражители делятся по признаку отношений между членами комплекса (рис. 3).
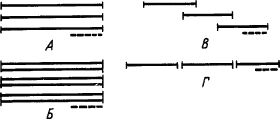
Рис. 3. Отношение во времени между членами комплексов сложных условных раздражителей. А — одновременный комплекс; Б — суммарный раздражитель; В — последовательный комплекс; Г — цепь раздражителей:
одинарными линиями показаны индифферентные раздражители, двойными — ранее выработанные сигналы, пунктиром — подкрепление
Условные рефлексы, выработанные на основе различных подкреплений. Основой для образования условного рефлекса — его подкреплением — может стать любая деятельность организма, осуществляемая нервной системой. Отсюда безграничные возможности условно-рефлекторной регуляции практически всех жизненных отправлений организма. На рис. 4 схематически представлены различные виды подкреплений, на базе которых можно выработать условные рефлексы.
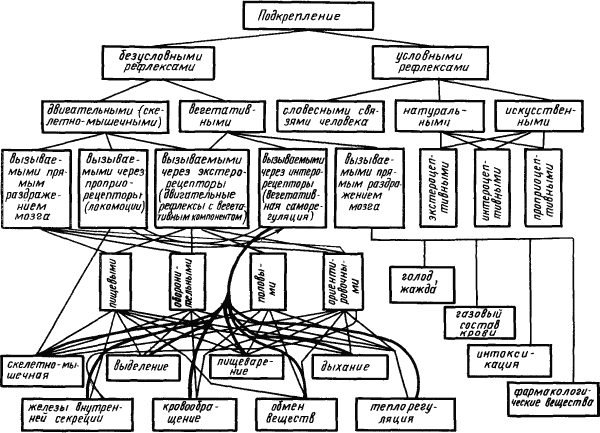
Рис. 4. Классификация подкреплений, на которые могут образовываться условные рефлексы
Каждый условный рефлекс, в свою очередь, может стать основой для образования нового условного рефлекса. Новая условная реакция, выработанная с помощью подкрепления сигнала другим условным рефлексом, называется условным рефлексом второго порядка. Условный рефлекс второго порядка, в свою очередь, можно использовать в качестве основы для выработки условного рефлекса третьего порядка и т.д.
Условные рефлексы второго, третьего и далее порядков широко распространены в природе. Они составляют наиболее значительную и совершенную часть натуральных условных рефлексов. Например, когда волчица кормит волчонка мясом растерзанной добычи, у него вырабатывается натуральный условный рефлекс первого порядка. Вид и запах мяса становятся для него сигналом еды. Затем он «учится» охоте. Теперь эти сигналы — вид и запах мяса пойманной добычи — играют роль основы для выработки охотничьих приемов подстерегания и преследования живой добычи. Так приобретают свое вторичное сигнальное значение разнообразные охотничьи признаки: обглоданный зайцем кустик, следы отбившейся от стада овцы и т.д. Они становятся раздражителями условных рефлексов второго порядка, выработанных на основе натуральных.
Наконец, исключительное разнообразие условных рефлексов, подкреплением которых служат другие условные рефлексы, встречается в высшей нервной деятельности человека. Более подробно они будут рассмотрены в гл. 17. Здесь лишь необходимо отметить, что в отличие от условных рефлексов животных условные рефлексы человека образуются не на основе безусловных пищевых, оборонительных и других подобных рефлексов, а на основе словесных сигналов, подкрепляемых результатами совместной деятельности людей. Поэтому мысли и поступки человека направляются не животными инстинктами, а мотивами его жизни в человеческом обществе.
Условные рефлексы, выработанные при разном соответствии во времени сигнала и подкрепления. По тому, как располагается во времени сигнал относительно подкрепляющей реакции, различают наличные и следовые условные рефлексы (рис. 5).
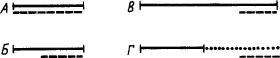
Рис. 5. Варианты временного соотношения сигнала и подкрепления. А — наличный совпадающий; Б — наличный отставленный; В — наличный запаздывающий; Г — следовой условной рефлекс:
сплошной линией обозначено время действия сигнала, прерывистой линией — время подкрепления
Наличными называют условные рефлексы, при выработке которых подкрепление применяют во время действия сигнального раздражителя. Наличные рефлексы разделяют в зависимости от срока присоединения подкрепления на совпадающие, отставленные и запаздывающие. Совпадающий рефлекс вырабатывается тогда, когда сразу после включения сигнала к нему присоединяют подкрепление. Например, при работе со слюнными рефлексами собаки включают звонок, а примерно через 1 с начинают собаку кормить. При таком способе выработки рефлекс образуется быстрее всего и скоро укрепляется.
Отставленный рефлекс вырабатывается в тех случаях, когда подкрепляющая реакция присоединяется лишь по истечении некоторого времени (до 30 с). Это наиболее употребительный способ выработки условных рефлексов, хотя он требует большего числа сочетаний, чем способ совпадения.
Запаздывающий рефлекс вырабатывается при присоединении подкрепляющей реакции после длительного изолированного действия сигнала. Обычно такое изолированное действие продолжается 1–3 мин. Этот способ выработки условного рефлекса еще труднее, чем оба предыдущих.
Следовыми называют условные рефлексы, при выработке которых подкрепляющую реакцию предъявляют лишь спустя некоторое время после выключения сигнала. В этом случае рефлекс вырабатывается на след от действия сигнального раздражителя; используют короткие интервалы (15–20 с) или длинные (1–5 мин). Образование условного рефлекса по следовому способу требует наибольшего числа сочетаний. Зато следовые условные рефлексы обеспечивают у животных весьма сложные акты приспособительного поведения. Примером может служить охота за скрывающейся добычей.
Условия выработки временных связей
Какие условия должны быть соблюдены, чтобы деятельность высших отделов центральной нервной системы могла завершаться выработкой условного рефлекса?
Сочетание сигнального раздражителя с подкреплением. Это условие выработки временных связей выявилось с первых же опытов со слюнными условными рефлексами. Шаги служителя, несущего пищу, только тогда вызывали «психическое слюноотделение», когда они сочетались с едой.
Этому не противоречит образование следовых условных рефлексов. Подкрепление сочетается в таком случае со следом возбуждения нервных клеток от ранее включенного и уже выключенного сигнала. Но если подкрепление станет опережать индифферентный раздражитель, то условный рефлекс удается выработать с большим трудом, лишь принимая ряд специальных мер. Это и понятно, так как если собаку сначала покормить, а потом дать сигнал еды, то, строго говоря, он даже не может называться сигналом, так как не предупреждает о предстоящих событиях, а отражает прошедшее. При этом безусловный рефлекс подавляет сигнальное возбуждение и препятствует образованию условного рефлекса на такой раздражитель.
Индифферентность сигнального раздражителя. Агент, выбранный в качестве условного раздражителя пищевого рефлекса, не должен сам по себе иметь какое-либо отношение к пище. Он должен быть безразличным, т.е. индифферентным, для слюнных желез. Сигнальный раздражитель не должен вызывать значительной ориентировочной реакции, мешающей образованию условного рефлекса. Однако ориентировочную реакцию вызывает каждый новый раздражитель. Поэтому для утраты им новизны его необходимо применять повторно. Лишь после того, как ориентировочная реакция будет практически погашена или сведена к незначительной величине, начинается образование условного рефлекса.
Преобладание силы возбуждения, вызванного подкреплением. Сочетание стука метронома и кормления собаки ведет к быстрому и легкому образованию у нее условного слюноотделительного рефлекса на этот звук. Но если пытаться сочетать с едой оглушительный стук механической трещотки, то такой рефлекс образовать чрезвычайно трудно. Для выработки временной связи большое значение имеет соотношение силы сигнала и подкрепляющей реакции. Чтобы между ними образовалась временная связь, очаг возбуждения, создаваемый последней, должен быть сильнее очага возбуждения, создаваемого условным раздражителем, т.е. должна возникать доминанта. Лишь тогда произойдет распространение возбуждения от очага индифферентного раздражителя к очагу возбуждения от подкрепляющего рефлекса.
Необходимость значительной интенсивности возбуждения именно подкрепляющей реакции имеет глубокий биологический смысл. В самом деле, условный рефлекс — это предупредительная реакция на сигнал о предстоящих существенных событиях. Но если раздражитель, который хотят сделать сигналом, оказывается событием даже более существенным, чем те, которые за ним последуют, то этот раздражитель сам вызывает соответствующую реакцию организма.
Отсутствие посторонних раздражителей. Каждое постороннее раздражение, например неожиданно донесшийся шум, вызывает уже упоминавшуюся ориентировочную реакцию. Собака настораживается, поворачивается в направлении звука и, главное, прекращает свою текущую деятельность. Животное все обращено навстречу новому раздражителю. Недаром И.П. Павлов называл ориентировочную реакцию рефлексом «Что такое?». Напрасно в это время экспериментатор будет подавать сигнал и предлагать собаке пищу. Условный рефлекс будет задержан более важным в данный момент для животного — ориентировочным рефлексом. Эта задержка создается дополнительным очагом возбуждения в коре мозга, который тормозит условное возбуждение и препятствует образованию временной связи. В природе множество таких случайностей влияет на ход образования условных рефлексов животных. Отвлекающая обстановка снижает продуктивность и умственной работы человека.
Нормальная работоспособность нервной системы. Полноценная замыкательная функция возможна при условии, если высшие отделы нервной системы находятся в нормальном рабочем состоянии. Метод хронического эксперимента потому и дал возможность обнаружить и изучить процессы высшей нервной деятельности, что при этом сохранялось нормальное состояние животного. Работоспособность нервных клеток мозга резко снижается при недостаточном питании, при действии ядовитых веществ, например бактериальных токсинов при болезнях, и т.д. Поэтому общее состояние здоровья является важным условием нормальной деятельности высших отделов мозга. Всем известно, как это условие влияет на умственную работу человека.
На образование условных рефлексов существенное влияние оказывает состояние организма. Так, физическая и умственная работа, условия питания, активность гормонов, действие фармакологических веществ, дыхание при повышенном или пониженном давлении, механические перегрузки и ионизирующее излучение в зависимости от интенсивности и сроков воздействия могут видоизменить, усилить или ослабить условно-рефлекторную деятельность вплоть до полного ее подавления.
Образование условных рефлексов и реализация актов высшей нервной деятельности в чрезвычайной степени зависят от потребности организма в биологически значимых агентах, используемых в качестве подкрепления. Так, у сытой собаки очень трудно выработать пищевой условный рефлекс, она будет отворачиваться от предлагаемой пищи, а у голодного животного с высокой пищевой возбудимостью он образуется быстро. Общеизвестно, как интерес учащегося к предмету занятий способствует лучшему его усвоению. Эти примеры показывают большое значение фактора отношения организма к проявляемым раздражителям, который обозначается как мотивация (К.В. Судаков, 1971).
Структурные основы замыкания временных условных связей
Исследование конечных, поведенческих проявлений высшей нервной деятельности значительно опередило изучение ее внутренних механизмов. До настоящего времени еще недостаточно изучены как структурные основы временной связи, так и ее физиологическая природа. По этому поводу высказываются разные взгляды, но вопрос еще не решен. Для его решения ведутся многочисленные исследования на системном и клеточном уровнях; используют электрофизиологические и биохимические показатели динамики функционального состояния нервных и глиальных клеток с учетом результатов раздражения или выключения различных структур мозга; привлекают данные клинических наблюдений. Однако на современном уровне исследований становится все более определенным, что при этом наряду со структурной необходимо учитывать и нейрохимическую организацию мозга.
Изменение локализации замыкания временных связей в эволюции. Независимо от того, считать ли, что условные реакции кишечнополостных (диффузная нервная система) возникают на основе суммационных явлений или настоящих временных связей, последние не имеют определенной локализации. У кольчатых червей (узловая нервная система) в опытах с выработкой условной реакции избегания было обнаружено, что при разрезании червя пополам рефлекс сохраняется у каждой половины. Следовательно, временные связи этого рефлекса замыкаются многократно, возможно, во всех нервных узлах цепочки и имеют множественную локализацию. У высших моллюсков (резко выражена анатомическая консолидация центральной нервной системы, образующей уже у осьминога развитый мозг) эксперименты с разрушением частей мозга показали, что надпищеводные отделы осуществляют многие условные рефлексы. Так, после удаления этих отделов осьминог перестает «узнавать» объекты своей охоты, теряет способность строить из камней убежище. У насекомых функции организации поведения сосредоточиваются в головных ганглиях. Особого развития у муравьев и пчел достигают так называемые грибовидные тела протоцеребрума, нервные клетки которых образуют множество синаптических контактов с многочисленными путями к другим отделам мозга. Предполагают, что именно здесь происходит замыкание временных связей при обучении насекомых.
Уже на раннем этапе эволюции позвоночных в передних отделах первоначально однородной мозговой трубки выделяется головной мозг, управляющий приспособительным поведением. В нем развиваются структуры, имеющие наибольшее значение для замыкания вредных связей в процессе условно-рефлекторной деятельности. На сновании опытов с удалением частей мозга у рыб было высказано предположение, что у них эту функцию выполняют структуры среднего и промежуточного мозга. Возможно, это определяется тем, что именно здесь сходятся пути всех сенсорных систем, а передний мозг развивается еще только как обонятельный.
У птиц ведущим отделом в развитии мозга становятся стриарные тела, образующие основную массу больших полушарий. Многочисленные факты указывают на то, что в них происходит замыкание временных связей. Голубь с удаленными полушариями служит наглядной иллюстрацией крайней бедности поведения, лишенного приобретенных в жизни навыков. Осуществление особенно сложных форм поведения птиц связывают с развитием структур гиперстриатума, образующих возвышение над полушариями, которое получило название «вульст». У врановых, например, его разрушение нарушает способность к осуществлению свойственных им сложных форм поведения.
У млекопитающих мозг развивается главным образом за счет бурного разрастания многослойной коры полушарий мозга. Особенное развитие получает новая кора (неокортекс), которая оттесняет старую и древнюю кору, покрывает в виде плаща весь мозг и, не умещаясь на его поверхности, собирается в складки, образуя многочисленные извилины, разделенные бороздами. Вопрос о структурах, осуществляющих замыкание временных связей, и их локализации в больших полушариях является предметом большого числа исследований и во многом является дискуссионным.
Удаление частей и всей коры больших полушарий. Если у взрослой собаки удалить затылочные области коры, то она утрачивает все сложные зрительные условные рефлексы и не может их восстановить. Такая собака не узнает своего хозяина, безразлична к виду самых лакомых кусков пищи, равнодушно смотрит на пробегающую мимо кошку, которую прежде бросилась бы преследовать. Наступает то, что раньше называли «психической слепотой». Собака видит, так как она обходит препятствия, поворачивается к свету. Но она «не понимает» смысла увиденного. Без участия зрительной коры зрительные сигналы остаются ни с чем не связанными.
И все-таки такая собака может образовывать очень простые зрительные условные рефлексы. Например, появление освещенной фигуры человека можно сделать сигналом пищи, вызывающим выделение слюны, облизывание, виляние хвостом. Следовательно, и в других областях коры находятся клетки, воспринимающие зрительные сигналы и способные их связывать с теми или иными действиями. Эти факты, подтвержденные и в опытах с повреждением корковых областей представительства других сенсорных систем, привели к мнению, что проекционные зоны перекрывают друг друга (Л. Лючиани, 1900). Дальнейшие исследования вопроса о локализации функций в коре в работах И.П. Павлова (1907–1909) показали широкое перекрытие проекционных зон, зависящее от природы сигналов и образуемых временных связей. Обобщая все эти исследования, И.П. Павлов (1927) выдвинул и обосновал представление о динамической локализации корковых функций. Перекрытия — это следы того широкого представительства всех видов рецепции во всей коре, которое имело место до их разделения по проекционным зонам. Каждое ядро корковой части анализатора окружено его рассеянными элементами, которых становится все меньше по мере удаления от ядра.
Рассеянные элементы не в состоянии заменить специализированные клетки ядра для образования тонких временных связей. Собака после удаления затылочных долей может вырабатывать лишь самые простые условные рефлексы, например, на вид освещенной фигуры. Заставить же ее различать две такие фигуры, сходные по форме, не удается. Однако если удаление затылочных долей произвести в раннем возрасте, когда еще не произошло выделение и закрепление проекционных зон, то, вырастая, эти животные обнаруживают способность к выработке сложных форм условных зрительных рефлексов.
Возможность широкой взаимозаменяемести функций коры мозга в раннем онтогенезе соответствует свойствам малодифференцированной коры мозга млекопитающих в филогенезе. С этой точки зрения получают объяснение результаты опытов на крысах, у которых степень нарушения условных рефлексов оказалась зависящей не от конкретной области удаляемой коры, а от общего объема удаляемой корковой массы (рис. 6). На основании этих опытов было сделано заключение, что для условно-рефлекторной деятельности все отделы коры имеют одинаковое значение, кора «эквипотенциальна» (К. Лешли, 1933). Однако результаты этих опытов могут лишь демонстрировать свойства малодифференцированной коры грызунов, а специализированная кора более высокоорганизованных животных обнаруживает не «эквипотенциальность», а хорошо выраженную динамическую специализацию функций.
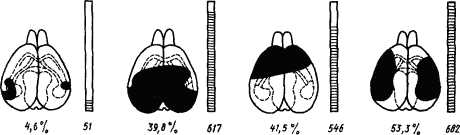
Рис. 6. Взаимозаменяемость частей коры мозга после их удаления у крыс (по К. Лешли):
удаленные области зачернены, цифрами под мозгом обозначена величина удаления в процентах ко всей поверхности коры, цифрами под столбиками — число ошибок при испытании в лабиринте
Первые опыты с удалением всей коры больших полушарий (<…пропуск…>Гольтц, 1982) показали, что после такой обширной операции, по-видимому, затрагивающей и ближайшую подкорку, собаки ничему не могли научиться. В опытах на собаках с удалением коры без травмирования подкорковых структур мозга удалось выработать простой условный рефлекс слюноотделения. Однако для его выработки потребовалось более 400 сочетаний, а угасить не удалось даже после 130 применений сигнала без подкрепления. Систематические исследования на кошках, которые легче, чем собаки, переносят операцию декортикации, показали трудность образования у них простых генерализованных пищевых и оборонительных условных рефлексов и выработки некоторых грубых дифференцировок. Опыты с холодовым выключением коры продемонстрировали, что полноценная целостная деятельность мозга без ее участия невозможна.
Разработка операции перерезки всех восходящих и нисходящих путей, связывающих кору с другими образованиями мозга, позволила осуществить декортикацию без прямого травмирования подкорковых структур и изучить роль коры в условно-рефлекторной деятельности. Оказалось, что у этих кошек можно было с большим трудом вырабатывать только грубые условные рефлексы общих движений, а оборонительного условного сгибания лапы не удалось получить даже после 150 сочетаний. Однако уже после 20 сочетаний на сигнал появилась реакция изменения дыхания и некоторые условные вегетативные реакции.
Конечно, при всех хирургических операциях трудно исключить их травмирующее влияние на подкорковые структуры и быть уверенным, что утраченная способность к тонкой условно-рефлекторной деятельности была функцией коры. Убедительные доказательства дали опыты с временным обратимым выключением функций коры, которое проявляется в распространяющейся депрессии электрической активности при аппликации KCI на ее поверхность. При выключении таким образом коры мозга крысы и испытании в это время реакции животного на условные и безусловные раздражители можно видеть, что безусловные рефлексы полностью сохраняются, а условные — нарушаются. Как видно из рис. 7, более сложные оборонительные и особенно пищевые условные рефлексы при максимальной депрессии в течение первого часа полностью отсутствуют, а простая защитная реакция избегания страдает в меньшей степени.
Таким образом, результаты экспериментов с частичной и полной хирургической и функциональной декортикацией говорят о том, что у высших животных функции формирования точных и тонких условных рефлексов, способных обеспечить приспособительное поведение, в основном выполняет кора больших полушарий.
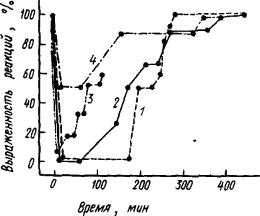
Рис. 7. Влияние временного выключения коры путем распространяющейся депрессии на пищевые (1) и оборонительные (2) условные рефлексы, безусловную реакцию избегания (3) и выраженность ЭЭГ (4) крысы (по Я. Бурешу и др.)
Корково-подкорковые отношения в процессах высшей нервной деятельности. Современные исследования подтверждают высказывание И.П. Павлова, что условно-рефлекторная деятельность осуществляется совместной работой коры и подкорковых структур. Из рассмотрения эволюции мозга как органа высшей нервной деятельности следует, что способность к образованию временных связей, обеспечивающих приспособительное поведение, проявляли структуры промежуточного мозга у рыб и стриарных (полосатых) тел у птиц, представляющие собой филогенетически наиболее молодые его отделы. Когда же у млекопитающих над этими отделами мозга возникла филогенетически самая молодая новая кора, осуществляющая наиболее тонкий анализ сигналов, то к ней перешла ведущая роль в образовании временных связей, организующих приспособительное поведение.
Структуры мозга, оказавшиеся подкорковыми, сохраняют в какой-то мере свои способности к замыканию временных связей, которые обеспечивают приспособительное поведение, свойственное тому уровню эволюции, когда эти структуры были ведущими. Об этом свидетельствует описанное выше поведение животных, которые после выключения коры мозга могли с трудом вырабатывать лишь очень примитивные условные рефлексы. Вместе с тем возможно, что такие примитивные временные связи не потеряли полностью своего значения и образуют часть нижнего уровня сложного иерархического механизма высшей нервной деятельности, возглавляемого корой больших полушарий.
Взаимодействие коры и подкорковых отделов мозга осуществляется и путем тонических влияний, регулирующих функциональное состояние нервных центров. Общеизвестно, как влияют настроение, эмоциональное состояние на эффективность умственной деятельности. И.П. Павлов говорил, что подкорка «заряжает» кору. Нейрофизиологические исследования механизмов подкорковых влияний на кору показали, что ретикулярная формация среднего мозга оказывает на нее восходящее активирующее действие. Получая коллатерали от всех афферентных путей, ретикулярная формация участвует во всех реакциях поведения, обусловливая деятельное состояние коры. Однако ее активирующее влияние при условном рефлексе организуется сигналами из проекционных зон коры (рис. 8). Раздражение ретикулярной формации вызывает изменение электроэнцефалограммы в виде ее десинхронизации, характерной для состояния активного бодрствования.
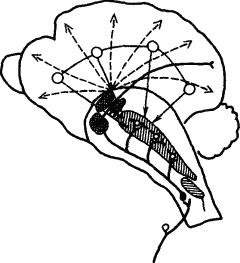
Рис. 8. Взаимодействие ретикулярной формации среднего мозга и коры (по Л.Г. Воронину):
жирными линиями обозначены афферентные специфические пути с коллатералями к ретикулярной формации, прерывистыми — восходящие пути к коре, тонкими — влияние коры на ретикулярную формацию, вертикальной штриховкой — облегчающая зона, горизонтальной — тормозная зона, клеточной штриховкой — таламические ядра
Иное действие на функциональное состояние коры оказывают специфические ядра таламуса. Их низкочастотное раздражение приводит к развитию в коре процессов торможения, которое может привести к засыпанию животного, и пр. Раздражение этих ядер вызывает в электроэнцефалограмме появление своеобразных волн — «веретен», которые переходят в медленные дельта-волны, характерные для сна. Ритм веретен может определяться тормозными постсинаптическими потенциалами (ТПСП) в нейронах гипоталамуса. Наряду с регуляторным влиянием неспецифических подкорковых структур на кору наблюдается и обратный процесс. Такие двусторонние корково-подкорковые взаимные влияния являются обязательными в осуществлении механизмов формирования временных связей.
Результаты некоторых экспериментов истолковывали как доказательство тормозящего действия стриарных структур на поведение животных. Однако дальнейшие исследования, в частности опыты с разрушением и стимуляцией хвостатых тел, и другие факты привели к заключению о наличии более сложных корково-подкорковых отношений.
Некоторые исследователи рассматривают факты об участии подкорковых структур в процессах высшей нервной деятельности как основание для того, чтобы считать их местом замыкания временных связей. Так возникло представление о «центрэнцефалической системе» как ведущей в поведении человека (У. Пенфилд, Г. Джаспер, 1958). В качестве доказательства замыкания временной связи в ретикулярной формации приводили наблюдения о том, что при выработке условного рефлекса первые изменения электрической активности мозга возникают именно в ретикулярной формации, а потом уже в коре больших полушарий. Но это свидетельствует лишь о вполне понятном раннем включении восходящей системы активации коры. Наконец, сильным аргументом в пользу подкорковой локализации замыкания считали возможность выработки условного, например зрительно-двигательного, рефлекса, несмотря на многократное рассечение коры на всю глубину, прерывающее все корковые пути между зрительной и двигательной областями. Однако этот экспериментальный факт не может служить доказательством, так как замыкание временной связи в коре имеет множественный характер и может происходить в любом ее участке между афферентными и эффекторными элементами. На рис. 9 жирными линиями показан путь условного зрительно-двигательного рефлекса при разрезах коры между зрительной и двигательной областями.
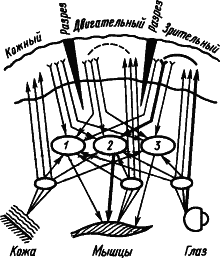
Рис. 9. Множественное замыкание временных связей в коре (показано пунктиром), которым не препятствуют ее разрезы (по А.Б. Когану):
1, 2, 3 — центральные механизмы оборонительных, пищевых и ориентировочных реакций соответственно; путь условного пищевого рефлекса на световой сигнал показан жирными линиями
Как показали многочисленные исследования, участие подкорковых структур в процессах высшей нервной деятельности не ограничивается регуляторной ролью ретикулярной формации среднего мозга и лимбических структур. Ведь уже на подкорковом уровне происходят анализ и синтез действующих раздражителей и оценка их биологического значения, в значительной мере определяющая характер образующихся связей с сигналом. Использование показателей формирования наиболее коротких путей, по которым сигнал достигает разных подкорковых структур мозга, выявило наиболее выраженное участие в процессах обучения задних отделов таламуса и поля CA3 гиппокампа. Роль гиппокампа в явлениях памяти подтверждается многими фактами. Наконец, нет оснований предполагать, что способность к примитивной замыкательной деятельности структур мозга, которая приобреталась в эволюции, когда они были ведущими, теперь у них полностью исчезла, когда эта функция перешла к новой коре.
Таким образом, корково-подкорковые отношения определяются регулированием функционального состояния коры активирующей системой — ретикулярной формацией среднего мозга и тормозной системой неспецифических ядер таламуса, а также возможным участием в образовании примитивных временных связей на нижнем уровне сложных иерархических механизмов высшей нервной деятельности.
Межполушарные отношения. Каким образом полушария головного мозга, являющиеся парным органом, участвуют в процессах образования условных связей? Ответ на этот вопрос был получен в опытах на животных, которым была сделана операция «расщепления» мозга путем перерезки мозолистого тела и передней комиссуры, а также продольного разделения перекреста зрительных нервов (рис. 10). После такой операции можно было вырабатывать разные условные рефлексы правого и левого полушарий, показывая разные фигуры правому или левому глазу. Если у оперированной таким образом обезьяны выработать условный рефлекс на световой стимул, подаваемый на один глаз, а затем подать его на другой глаз, то никакой реакции не последует. «Обучение» одного полушария оставляло другое «необученным». Однако при сохранении мозолистого тела и другое полушарие оказывается «обученным». Мозолистое тело осуществляет межполушарный перенос навыка.
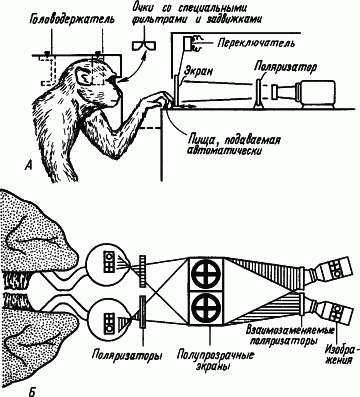
Рис. 10. Исследования процессов обучения у обезьян, подвергнутых операции «расщепления» мозга. А — устройство, направляющее в правый глаз одно изображение, а в левый — другое; Б — специальная оптика для проецирования зрительных образов в разные глаза (по Р. Сперри)
С помощью метода функционального выключения коры мозга у крыс воспроизводили на некоторое время условия «расщепленного» мозга. При этом временные связи могло образовать одно оставшееся активным полушарие. Этот рефлекс проявлялся и после прекращения действия распространяющейся депрессии. Он сохранился даже после инактивации полушария, которое было активным во время выработки этого рефлекса. Следовательно, «обученное» полушарие передало по волокнам мозолистого тела приобретенный навык «необученному». Однако этот рефлекс исчезал, если такую инактивацию производили до того, как полностью восстановится активность полушария, включенного во время выработки условного рефлекса. Таким образом, для переноса приобретенного навыка из одного полушария в другое необходимо, чтобы оба они были активны.
Дальнейшие исследования межполушарных отношений при образовании временных связей условных рефлексов показали, что во взаимодействии полушарий специфическую роль играют процессы торможения. Так, полушарие, противоположное стороне подкрепления, становится доминирующим. Оно вначале осуществляет формирование приобретенного навыка и его перенос в другое полушарие, а потом, притормаживая активность противоположного полушария и оказывая избирательно тормозное влияние на структуру временных связей, совершенствует условный рефлекс.
Таким образом, каждое полушарие, даже будучи изолированным от другого, способно к образованию временных связей. Однако в естественных условиях их парной работы сторона подкрепления определяет доминантное полушарие, которое формирует тонкую возбудительно-тормозную организацию условно-рефлекторного механизма приспособительного поведения.
Предположения о месте замыкания временных связей в больших полушариях головного мозга. Открыв условный рефлекс, И.П. Павлов сначала предположил, что временная связь является «вертикальной связью» между зрительной, слуховой или иными частями коры больших полушарий и подкорковыми центрами безусловных рефлексов, например пищевого — корково-подкорковая временная связь (рис. 11, А). Однако многочисленные факты дальнейшей работы и результаты специальных опытов привели затем к выводу, что временная связь — это «горизонтальная связь» между очагами возбуждения, находящимися в пределах коры. Например, во время образования условного слюнного рефлекса на звук звонка замыкание происходит между клетками слухового анализатора и клетками, которые представляют в коре безусловный слюноотделительный рефлекс (рис. 11, Б). Такие клетки назвали представителями безусловного рефлекса.
Наличие в коре больших полушарий собаки представительства безусловных рефлексов доказывается следующими фактами. Если применять в качестве пищевого раздражителя сахар, то слюноотделение на него вырабатывается лишь постепенно. Если какой-либо условный раздражитель не подкреплять, то следующее за ним «сахарное» слюноотделение уменьшается. Значит, этот безусловный рефлекс имеет нервные клетки, расположенные в сфере протекания корковых процессов. Дальнейшие исследования показали, что если удалить у собаки кору, то ее безусловные рефлексы (слюнные, отделение желудочного сока, движения конечностей) претерпевают стойкие изменения. Следовательно, безусловные рефлексы кроме подкоркового центра имеют центры и на корковом уровне. Вместе с тем раздражитель, который сделан условным, также имеет представительство в коре. Отсюда возникло предположение (Э.А. Асратян, 1963), что временные связи условного рефлекса замыкаются между этими представительствами (рис. 11, В).
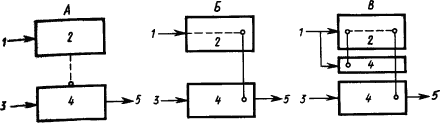
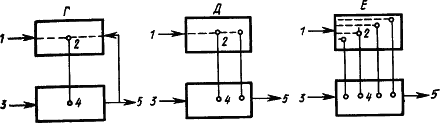
Рис. 11. Различные предположения о структуре временной связи условного рефлекса (объяснение см. в тексте):
1 — условный раздражитель, 2 — корковые структуры, 3 — безусловный раздражитель, 4 — подкорковые структуры, 5 — рефлекторная реакция; прерывистыми линиями показаны временные связи
Рассмотрение процессов замыкания временных связей как центральных звеньев формирования функциональной системы (П.К. Анохин, 1961) относит замыкание к структурам коры, где происходит сопоставление содержания сигнала — афферентный синтез — и результата условно-рефлекторного ответа — акцептор действия (рис. 11, Г).
Изучение двигательных условных рефлексов показало сложную структуру образующихся при этом временных связей (Л.Г. Воронин, 1952). Каждое совершающееся по сигналу движение само становится сигнальным для складывающихся при этом двигательных координаций. Образуются две системы временных связей: на сигнал и на движение (рис. 11, Д).
Наконец, исходя из того, что условные рефлексы сохраняются при хирургическом разделении сенсорных и моторных корковых областей и даже после множественных разрезов коры, а также учитывая, что кора обильно снабжена как входящими, так и выходящими путями, было высказано предположение, что замыкание временных связей может происходить в каждом ее микроучастке между его афферентными и эфферентными элементами, которые активируют центры соответствующих безусловных рефлексов, служащих подкреплением (А.Б. Коган, 1961) (см. рис. 9 и 11, Е). Такое предположение соответствует представлению о возникновении временной связи в пределах анализатора условного раздражителя (О.С. Адрианов, 1953), мнению о возможности «местных» условных рефлексов, замыкающихся внутри проекционных зон (Э.А. Асратян, 1965, 1971), и заключению, что в замыкании временной связи ключевую роль играет всегда афферентное звено (У.Г. Гасанов, 1972).
Нейронная структура временной связи в коре больших полушарий. Современные сведения о микроскопическом строении коры больших полушарий в сочетании с результатами электрофизиологических исследований позволяют с известной степенью вероятности судить о возможном участии тех или иных корковых нейронов в образовании временных связей.
Высокоразвитая кора мозга млекопитающих, как известно, делится на шесть слоев разного клеточного состава. Приходящие сюда нервные волокна заканчиваются большей частью у клеток двух видов. Один из них — это вставочные нейроны, расположенные во II, III и отчасти IV слоях. Их аксоны идут в V и VI слои к крупным пирамидным клеткам ассоциативного и центробежного типа. Это наиболее короткие пути, которые, возможно, и представляют собой врожденные связи корковых рефлексов.
Другой вид клеток, с которыми приходящие волокна образуют наибольшее число контактов, составляют кустовидно ветвящиеся округлые и угловатые короткоотростчатые клетки, часто имеющие звездчатую форму. Они располагаются главным образом в IV слое. Их количество увеличивается с развитием мозга млекопитающих. Это обстоятельство наряду с тем, что звездчатые клетки занимают положение конечной станции для приходящих в кору импульсов, позволяет предположить, что именно звездчатые клетки являются основными воспринимающими корковыми клетками анализаторов и что увеличение их численности в эволюции представляет собой морфологическую основу достижения высокой тонкости и точности отражения окружающего мира.
Система вставочных и звездчатых нейронов может вступать в бесчисленное множество контактов с ассоциативными и проекционными крупными нейронами пирамидной формы, расположенными в V и VI слоях. Ассоциативные нейроны своими аксонами, проходящими через белое вещество, соединяют между собой разные корковые ноля, а проекционные нейроны дают начало путям, соединяющим кору с низшими отделами мозга.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава II С чего начинается развитие. Образование яйца
Глава II С чего начинается развитие. Образование яйца К вопросу о том, с чего начинается развитие, не следует относиться слишком серьезно. Обычно в литературе началом эмбрионального развития считают оплодотворение. Однако если видеть в оплодотворении некий символ
Глава 5 Земледелие и образование городов. Стресс у мышей и людей — и у шимпанзе.
Глава 5 Земледелие и образование городов. Стресс у мышей и людей — и у шимпанзе. Земледелие влекло за собой не только рост достатка, оно изменило образ жизни человека как в материальном, так и в социальном отношении.Хорошие урожаи позволяли большему числу людей постоянно
6.3. Торможение условных рефлексов
6.3. Торможение условных рефлексов И.П. Павлов выделил две разновидности торможения условных рефлексов – безусловное (внешнее) и условное (внутреннее) торможение.Безусловное торможение. Полная остановка начавшегося рефлекса или снижение его активности под воздействием
ГЛАВА 2 Образование нашей планеты: «холодная» и «горячая» гипотезы. Гравитационная дифференциация недр. Происхождение атмосферы и гидросферы
ГЛАВА 2 Образование нашей планеты: «холодная» и «горячая» гипотезы. Гравитационная дифференциация недр. Происхождение атмосферы и гидросферы Рассказ о происхождении Земли и Солнечной системы нам придется начать издалека. В 1687 году И. Ньютон вывел закон всемирного
Глава 4 ТОРМОЖЕНИЕ УСЛОВНЫХ РЕФЛЕКСОВ
Глава 4 ТОРМОЖЕНИЕ УСЛОВНЫХ РЕФЛЕКСОВ До сих пор был рассмотрен лишь один основной физиологический процесс высшей нервной деятельности: процесс возбуждения нейронов и его влияние на выработку условных рефлексов. Другим основным физиологическим процессом высшей
НАУЧЕНИЕ В ФОРМЕ КЛАССИЧЕСКИХ УСЛОВНЫХ РЕФЛЕКСОВ
НАУЧЕНИЕ В ФОРМЕ КЛАССИЧЕСКИХ УСЛОВНЫХ РЕФЛЕКСОВ Немного историиПочему условный рефлекс во всех наших книгах о дрессировке? Потому, что только он и ничего другого? Потому, что в научении и поведении он преобладает? Потому, что все можно свести к условному рефлексу? Да нет,
НАУЧЕНИЕ НА ОСНОВЕ ОБРАЗОВАНИЯ ИНСТРУМЕНТАЛЬНЫХ УСЛОВНЫХ РЕФЛЕКСОВ (ОПЕРАНТНОЕ НАУЧЕНИЕ)
НАУЧЕНИЕ НА ОСНОВЕ ОБРАЗОВАНИЯ ИНСТРУМЕНТАЛЬНЫХ УСЛОВНЫХ РЕФЛЕКСОВ (ОПЕРАНТНОЕ НАУЧЕНИЕ) Отцом инструментальной формы научения считается Э. Торндайк, который еще в конце прошлого века назвал такую форму научения обучением «методом проб, ошибок и случайного успеха».
Глава 24. Образование и обезвреживание NH3 в организме
Глава 24. Образование и обезвреживание NH3 в организме В состоянии азотистого равновесия организм взрослого человека потребляет и выделяет около 15 г азота за сутки. Из экскретируемого с мочой азота на долю мочевины приходится 85%, креатинина – 5%, аммонийных солей – 3%,
3.4.1. Приспособительные механизмы, отличные от условных рефлексов
3.4.1. Приспособительные механизмы, отличные от условных рефлексов Для обучения характерно изменение чувствительности к действию определенных стимулов и способности на них реагировать Простейший вид обучения — суммационная реакция, описанная еще И.М. Сеченовым. Она
Правомерно ли сводить поведение собаки к простому комплексу условных и безусловных рефлексов?
Правомерно ли сводить поведение собаки к простому комплексу условных и безусловных рефлексов? В 2001 году опубликованы результаты эксперимента, проведенного сотрудниками Института эволюционной антропологии в Лейпциге (Германия). Изучая зависимость поведения собак от
ГЛАВА 3 ОБРАЗОВАНИЕ ПОЛОВЫХ КЛЕТОК
ГЛАВА 3 ОБРАЗОВАНИЕ ПОЛОВЫХ КЛЕТОК Размножение становится возможным лишь по достижении животным половой зрелости. Половая зрелость животных наступает с момента начала активного образования половых клеток. Сроки наступления ее у собак сильно варьируют и зависят от
8.4. Образование человеческих рас
8.4. Образование человеческих рас Процесс дифференциации вида Homo sapiens и образования рас также является весьма дискуссионным разделом антропологии. На протяжении всей истории антропологии шел спор между сторонниками теорий моноцентризма и полицентризма. Он то
Классическая выработка условных рефлексов
Классическая выработка условных рефлексов Классическая выработка условных рефлексов заключается в сочетании стимула, который производит некоторую реакцию, с другим, нейтральным. Затем, когда действует только второй стимул, он пробуждает ту же реакцию».Под «методами
[ВЛИЯНИЕ ИДЕАЛИСТИЧЕСКОГО МИРОВОЗЗРЕНИЯ НЛ ОТНОШЕНИЕ УЧЕНЫХ К УЧЕНИЮ ОБ УСЛОВНЫХ РЕФЛЕКСАХ] [ 67 ]
[ВЛИЯНИЕ ИДЕАЛИСТИЧЕСКОГО МИРОВОЗЗРЕНИЯ НЛ ОТНОШЕНИЕ УЧЕНЫХ К УЧЕНИЮ ОБ УСЛОВНЫХ РЕФЛЕКСАХ][ 67 ] Акад. И. П. Павлов. — ...Вы знаете, как я сообщал из истории развития учения о наших условных рефлексах — наши условные рефлексы встречают сильное препятствие в головах у людей,
4. Образование
4. Образование Вторым средовым фактором, который иногда считают ответственным за низкие величины IQ народов экономически развивающихся стран, является недостаток образования. К примеру, Фиш (Fish; 2002, с. 14) пишет: «Недостаток систематического образования африканцев