Глава 12 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЛИЧИНОЧНО-ХОРДОВЫХ КРУГЛОРОТЫХ И РЫБ
Глава 12
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЛИЧИНОЧНО-ХОРДОВЫХ КРУГЛОРОТЫХ И РЫБ
Высшая нервная деятельность позвоночных животных отражает одну из важных тенденций их эволюции — индивидуальное совершенствование. Эта тенденция проявляется в нарастающей продолжительности жизни, сокращении численности потомства, увеличении размеров тела, усилении консерватизма наследственности. Выражением этой же тенденции является и то, что на основе ограниченного числа видовых инстинктов каждый индивидуум в порядке личного жизненного опыта может образовывать большее количество разнообразнейших условных рефлексов.
У таких низших хордовых животных, как личиночно-хордовые и круглоротые, условные рефлексы имеют примитивный характер. С развитием аналитико-синтетической деятельности мозга и использованием все более тонких сигналов у рыб условные рефлексы начинают играть все более значительную роль в их поведении.
Условные рефлексы личиночно-хордовых
Несмотря на регресс своей нервной системы, асцидия может образовать условный защитный рефлекс закрытия сифонов на звуковой, или, вернее, вибрационно-механический сигнал.
Для выработки такого рефлекса над сидящей в аквариуме асцидией устанавливали капельницу. При каждом ударе капли о поверхность воды асцидия быстро закрывала сифоны, а при более сильном раздражении (падение капли с большой высоты) втягивала их внутрь. Источником условных сигналов служил электрический звонок, укрепленный на столе рядом с аквариумом. Его изолированное действие продолжалось 5 с, в конце которых падала капля. После 20–30 сочетаний сам звонок мог уже вызывать защитные движения сифонов.
Удаление центрального нервного узла уничтожало выработанный рефлекс и делало невозможным образование новых. Настойчивые попытки выработать у здоровых животных аналогичные условные рефлексы на свет оказались безуспешными. Очевидно, отсутствие реакций на световые сигналы объясняется условиями жизни асцидий.
В этих опытах также обнаружилось, что в результате сочетаний сигнала с безусловной реакцией последняя все легче вызывалась безусловным раздражителем. Возможно, что такое условное повышение возбудимости сигнализируемой реакции представляет собой начальную суммационную форму временной связи, из которой потом развивались более специализированные.
Круглоротые
Морская минога достигает метра в длину. Половой инстинкт каждую весну заставляет ее так же, как многих морских рыб, покидать глубины моря и подниматься в реки для икрометания. Однако на эту инстинктивную реакцию может быть выработано торможение (миноги перестали входить в реки, где они встречали загрязненную воду).
Условные рефлексы речной миноги исследовали при подкреплении ударами электрического тока. Световой сигнал (2 лампы по 100 Вт), к которому через 5–10 с изолированного действия присоединяли 1–2-секундное безусловное электрокожное раздражение, уже через 3–4 сочетания сам начинал вызывать двигательную оборонительную реакцию. Однако после 4–5 повторений условный рефлекс уменьшался и вскоре исчезал. Через 2–3 ч его можно было вырабатывать заново. Примечательно, что одновременно с уменьшением условного оборонительного рефлекса уменьшалась величина и безусловного. Порог электрокожного раздражения для вызова оборонительной реакции при этом повышался. Возможно, что такие изменения зависели от травмирующего характера электрического раздражения.
Как было показано выше на примере асцидий, образование условного рефлекса может проявляться в повышении возбудимости сигнализируемой реакции. В данном случае на примере миноги видно, как при торможении условного рефлекса возбудитель сигнализируемой реакции снижается. С легкостью образуя условный оборонительный рефлекс на свет лампы, миноги оказались не в состоянии выработать его на звук звонка. Несмотря на 30–70 сочетаний звонка с электрическими ударами, он так и не стал сигналом защитных движений. Это свидетельствует о преимущественно зрительной ориентировке миног в окружающей обстановке.
Минога воспринимает световые раздражения не только при помощи глаз. Даже после перерезки зрительных нервов или полного удаления глаз реакция на свет сохранялась. Она исчезала только тогда, когда кроме глаза был удален и теменной орган головного мозга, имеющий светочувствительные клетки. Фоторецепторной функцией обладают также некоторые нервные клетки промежуточного мозга и клетки расположенные в коже около анального плавника.
Рыбы
Достигнув высокого совершенства в приспособлении к водному образу жизни, рыбы значительно расширили свои рецепторные возможности, в частности, за счет механорецепторов органов боковой линии. Условные рефлексы составляют существенную часть поведения хрящевых и особенно костистых рыб.
Хрящевые рыбы. Прожорливость акулы недаром вошла в поговорку. Ее мощный пищевой инстинкт трудно затормозить даже сильными болевыми раздражениями. Так, китобои утверждают, что акула продолжает рвать и глотать куски мяса убитого кита, даже если вонзить в нее острогу. На основе столь резко выраженных безусловных пищевых реакций у акул в природной обстановке, по-видимому, образуются многие условные пищевые рефлексы. Об этом, в частности, свидетельствуют описания того, как быстро акулы вырабатывают реакцию сопровождения кораблей и даже подплывают в определенное время к борту, с которого выбрасываются кухонные отходы.
Акулы очень активно используют обонятельные сигналы пищи. Известно, как они преследуют раненую добычу по следу крови. Значение обоняния для образования пищевых рефлексов было показано в опытах на небольших Mustelus laevis, свободно плавающих в пруду. Эти акулы находили живых спрятавшихся крабов за 10–15 мин, а убитых и вскрытых — через 2–5 мин. Если акулам закрывали ноздри ватой с вазелином, то они не могли найти спрятавшегося краба.
Свойства образования условных оборонительных рефлексов у черноморских акул (Squalus acanthias) изучали с помощью методики, описанной выше для миног. Оказалось, что акулы вырабатывали условный рефлекс на звонок после 5–8 сочетаний, а на лампу — лишь после 8–12 сочетаний. Выработанные рефлексы были очень нестойкими. Они не сохранялись в течение суток, и на следующий день их нужно было вырабатывать вновь, хотя для этого требовалось меньше сочетаний, чем в первый день.
Сходные свойства образования условных оборонительных рефлексов обнаружили и другие представители хрящевых рыб — скаты. В этих свойствах находят свое отражение условия их жизни. Так, обитателю морских глубин шиповатому скату для выработки рефлекса на звонок понадобились 28–30 сочетаний, в то время как подвижному жителю прибрежных вод скату-хвостоколу было достаточно 4–5 сочетаний. В этих условных рефлексах также проявилась непрочность временных связей. Выработанный накануне условный рефлекс на следующий день исчезал. Его нужно было каждый раз восстанавливать двумя-тремя сочетаниями.
Костистые рыбы. Благодаря громадному разнообразию в строении тела и поведении, костистые рыбы достигли отличной приспособленности к самым различным условиям обитания. К этим рыбам принадлежит и малютка Mistichthus luzonensis (самое маленькое позвоночное, размером в 12–14 мм), и гигантский «сельдяной король» (Regalecus) южных морей, достигающий 7 м в длину.
Чрезвычайно разнообразны и специализированы инстинкты рыб, особенно пищевые и половые. Одни рыбы, как, например, вегетарианец карась, мирно плавают в илистых водоемах, другие, как, например, плотоядная щука, живут охотой. Хотя большинство рыб оставляет оплодотворенную икру на произвол судьбы, некоторые из них проявляют заботу о потомстве. Так, морские собачки охраняют отложенные яйца до вылупления молоди. Девятииглая колюшка строит настоящее гнездо из травинок, склеивая их своими слизистыми выделениями. Закончив постройку, самец загоняет в гнездо самку и не выпускает, пока она не вымечет икру. После этого он поливает икру семенной жидкостью и сторожит у входа в гнездо, время от времени вентилируя его особыми движениями грудных плавников.
Пресноводные рыбы из семейства Cichlidae в случае опасности прячут вылупившуюся молодь в рот. Описывают особые «зовущие» движения взрослых рыб, которыми они собирают своих мальков. Пинагор водит за собой мальков, которые могут прикрепляться к телу отца особыми присосками.
Ярким проявлением силы полового инстинкта рыб являются сезонные миграции. Например, лососевые в определенное время года устремляются из моря в реки для нерестования. Их массами истребляют звери и птицы, множество рыб гибнет от истощения, но оставшиеся упорно продолжают свой путь. В неудержимом стремлении к верховьям реки благородный лосось, встречая препятствие, прыгает на камни, разбивается в кровь и вновь бросается вперед, пока его не преодолеет. Он перескакивает пороги и взбирается на водопады. Защитный и пищевой инстинкты полностью затормаживаются, все подчинено задаче размножения.
Взаимоотношения рыб в стае обнаруживают определенную иерархию подчинения вожаку, которая может принимать различные формы. Так, приводят наблюдения за стайкой рыбок малабарских данио, где вожак плывет почти горизонтально, что позволяет ему первому увидеть и схватить упавшее на поверхность воды насекомое. Остальные рыбки распределяются по рангам и плывут с наклоном от 20 до 45°. Большую роль в поведении рыб играют выделяемые ими феромоны. Например, при повреждении кожи пескаря в воду поступают торибоны — химические сигналы тревоги. Достаточно было капнуть такой воды в аквариум с пескарями, чтобы они бросились в бегство.
Условные рефлексы на звуковые раздражители. Любители аквариумов хорошо знают, как можно приучить рыбок собираться у поверхности воды на сигнал постукивания по стенке, если практиковать такое постукивание перед каждым кормлением. По-видимому, подобный условный пищевой рефлекс определял поведение знаменитых рыб монастырского пруда в Кремсе (Австрия), привлекающих внимание туристов тем, что они приплывали к берегу на звук колокольчика. Исследователи, отрицающие слух у рыб, утверждают, что рыбы приплывали лишь тогда, когда они видели приходящего к пруду человека или когда его шаги вызывали сотрясение почвы. Однако этим не исключается участие звука в качестве одной из частей комплексного раздражителя.
Вопрос о слухе рыб долгое время оставался спорным, тем более, что рыба не имеет ни улитки, ни основной мембраны кортиева органа. Он был решен положительно лишь объективным методом условных рефлексов (Ю. Фролов, 1925).
Опыты были поставлены на пресноводных (карась, ерш) и морских (треска, пикша, бычок) рыбах. В небольшом аквариуме испытуемая рыба плавала на поводке-нитке, привязанном к капсуле воздушной передачи. Эту же нитку использовали для подведения к телу рыбы электрического тока, вторым полюсом служила лежащая на дне металлическая пластинка. Источником звука являлась телефонная трубка. После 30–40 сочетаний звуков с электрическими ударами образовывался слуховой условный защитный рефлекс. При включении телефона рыба ныряла, не ожидая электрического удара.
Таким способом можно было выработать условные рефлексы также на различного рода вибрации воды и другие сигналы, как, например, свет.
Выработанные на подкреплении электрическим током оборонительные рефлексы оказались очень прочными. Они сохранялись в течение длительного времени и с трудом поддавались угашению. Вместе с тем на следы сигналов рефлексы выработать не удалось. Если начало безусловного подкрепления отставало от конца действия условного сигнала хотя бы на 1 с, рефлекс не образовывался. Обнаружили также, что выработка одного условного рефлекса облегчала образование последующих. По результатам этих опытов можно судить о некоторой инертности и слабости временных связей, способных, однако, к тренировке.
Условный пищевой рефлекс на звук нетрудно выработать у золотистой рыбки орфы, сопровождая звуковой сигнал опусканием в аквариум мешочка с нарезанными червями. У рыбки Umbra limi был не только образован подобный условный положительный рефлекс на тон 288 колебаний/с, но и выработано дифференцирование тона 426 колебаний/с, который сопровождался подачей вместо пищи комочка фильтровальной бумаги, смоченной камфорным спиртом.
Чтобы полностью исключить участие зрения, звуковые условные рефлексы вырабатывали на предварительно ослепленных карликовых сомиках, гольянах и гольцах. Этим способом была установлена верхняя граница слышимости звуков, которая оказалась у сомика около 12 000 колебаний/с, у гольяна — около 6000, у гольца — около 2500. При определении нижней границы слышимости звуков, оказалось, что рыбы воспринимают очень медленные (2–5 колебаний/с) и даже одиночные колебания воды, которые для человеческого уха не являются звуками. Эти медленные колебания можно сделать условными раздражителями пищевого рефлекса и выработать их дифференцирование. Перерезка нервов органа боковой линии уничтожает рефлексы на низкие звуки, нижний предел слышимости поднимается до 25 Гц. Следовательно, орган боковой линии является своеобразным органом инфразвукового слуха рыб.
За последнее время накоплены сведения о звуках, издаваемых рыбами. Давно известно, что малайские рыбаки ныряют в воду, чтобы по слуху узнать, где находится косяк рыбы. «Голоса» рыб записывают на пленку магнитофона. Они оказались разными у различных видов рыб, более высокими у мальков и более низкими у взрослых. Среди наших черноморских рыб наиболее «голосистым» оказался горбыль. Примечательно, что у горбыля условный рефлекс на звук образуется после 3–5 сочетаний, т.е. быстрее, чем у других исследованных рыб, например у карася, потребовавшего 9–15 сочетаний. Однако на световые сигналы горбыль вырабатывает условные рефлексы хуже (после 6–18 сочетаний) .
Условные рефлексы на световые раздражители. Разнообразные условные рефлексы на пищевом подкреплении вырабатывали при дрессировках рыб с целью изучить их зрение. Так, в опытах с гольянами установлено, что они хорошо дифференцируют световые раздражения по яркости, различая разные оттенки серого, удалось также добиться различения рыбами штриховых фигур, Причем вертикальная штриховка приобрела сигнальное значение быстрее, чем горизонтальная. Опыты с окунями, пескарями и гольянами показали, что рыбы могут вырабатывать дифференцировки по форме таких фигур, как треугольник и квадрат, круг и овал. Оказалось также, что рыбам свойственны зрительные контрасты, отражающие индукционные явления в мозговых частях анализаторов.
Если кормить макроподов красными личинками хирономид, то вскоре рыбки набрасывались на стенку аквариума, когда снаружи к стеклу приклеивали комочки красной шерсти, сходные по размерам с личинками. На зеленые и белые комочки тех же размеров микроподы не реагировали. Если же кормить рыбок катышками белого хлебного мякиша, то они начинают хватать оказавшиеся в поле зрения белые шерстяные комочки.
Описывают, что однажды коралловому хищнику дали выкрашенную в красный цвет атеринку вместе с щупальцем медузы. Хищная рыба сначала схватила добычу, но, обжегшись о стрекательные капсулы, немедленно ее выпустила. После этого она не брала красных рыбок в течение 20 дней.
Особенно много исследований проведено по изучению свойств зрения карпов. Так, в опытах выработки оборонительных условных рефлексов на предъявление линий в качестве сигналов показано, что рыбы могли дифференцировать их по углу наклона. На основании этих и других экспериментов высказаны предположения о возможном механизме зрительного анализа у рыб с помощью детекторных нейронов. О высоком развитии зрительного восприятия карпа свидетельствует его способность различать краску предмета даже в разных условиях освещения. Такое свойство константности восприятия проявилось у карпа и в отношении формы предмета, реакция на который оставалась определенной, несмотря на его пространственные преобразования.
Условные обонятельные, вкусовые и температурные рефлексы. Рыбы могут вырабатывать обонятельные и вкусовые условные рефлексы. После того как гольяна некоторое время кормили мясом с запахом мускуса, он начал реагировать типичной поисковой реакцией на ранее безразличный для него мускусный запах. Обонятельным сигналом можно было сделать запах скатола или кумарина. Сигнальный запах дифференцировался от неподкрепляемых кормлением. Очень легко становится положительным сигналом для гольянов запах слизи, покрывающей их тело. Возможно, что таким натуральным рефлексом объясняются некоторые свойства стадного поведения этих рыб.
Если дождевых червей, скармливаемых гольянам, предварительно вымачивать в сахарном растворе, то через 12–14 дней рыбы будут набрасываться на опущенную в аквариум вату с раствором сахара. Такую же реакцию вызывали и другие сладкие вещества, в том числе сахарин и глицерин. Можно выработать вкусовые условные рефлексы на горькое, соленое, кислое. Порог раздражения горьким оказался у гольяна выше, а сладким ниже, чем у человека. Эти рефлексы не зависели от запаховых сигналов, так как они сохранялись и после удаления обонятельных долей мозга.
Описаны наблюдения, показавшие, что развитие у рыб хеморецепторов связано с поиском и обнаружением пищи. У карпов можно выработать инструментальные условные рефлексы регулирования солености или кислотности воды. При этом двигательная реакция приводила к добавлению растворов заданной концентрации. У рыбок Poecilia reticulata Peters вырабатывали условные пищевые рефлексы на вкус бета-фенилэтанола с дифференцировкой на кумарин.
Получены убедительные доказательства того, что лососевые, приблизившись к устью реки, где они родились, пользуются обонянием, чтобы найти свое «родное» нерестилище. На высокую избирательную чувствительность их хеморецепции указывают результаты электрофизиологического опыта, в котором импульсы регистрировались в обонятельной луковице, только когда через ноздри рыбы пропускали воду из «родного» нерестилища, и отсутствовали, если вода была из «чужого». Известно использование форели в качестве тест-объекта для оценки чистоты воды после очистных сооружений.
Можно сделать температуру воды, в которой плавает рыба, условным пищевым сигналом. При этом удалось добиться дифференцирования температурных раздражителей с точностью до 0,4 °C. Есть основания полагать, что натуральные температурные сигналы играют большую роль в половом поведении рыб, в частности в нерестовых миграциях.
Сложные пищедобывательные рефлексы. Для лучшего сравнения показателей условно-рефлекторной деятельности разных видов животных используют естественные пищедобывательные движения. Таким движением для рыб является схватывание подвешенной на нитке бусинки. Первые случайные схватывания подкрепляют кормом и сочетают со слуховым или зрительным сигналом, на который образуется условный рефлекс. Такой условный зрительный рефлекс, например, был образован и укреплен у карася за 30–40 сочетаний. Также была выработана дифференцировка по цвету и условный тормоз. Однако повторные переделки сигнального значения положительного и отрицательного раздражителей оказались чрезвычайно трудной задачей для рыб и даже приводили к расстройствам условно-рефлекторной деятельности.
Исследования поведения рыб в лабиринтах показали их способность к выработке реакции безошибочного выбора правильного пути.
Так, темнолюбивая рыбка Tundulus после 12–16 проб в течение двух дней стала проплывать через отверстия ширм, не заходя в тупики, прямо в угол, где ждала пища. В аналогичных опытах с золотыми рыбками время поисков выхода из лабиринта за 36 проб уменьшилось со 105 до 5 мин. После 2 недель перерыва в работе приобретенный навык изменился лишь немного. Однако с более сложными лабиринтами, типа применявшихся для крыс, рыбы не могли справиться, несмотря на сотни проб.
У хищных рыб можно выработать условно-рефлекторное подавление охотничьего инстинкта.
Если в аквариум со щукой поместить за стеклянной перегородкой карася, то щука сразу бросится на него. Однако после нескольких ударов головой головой в стекло нападения прекращаются. Через несколько дней щука уже не пытается схватить карася. Натуральный пищевой рефлекс полностью угашен. Тогда снимают перегородку, и карась может плавать рядом со щукой. Аналогичный опыт был проведен с хищными окунями и пескарями. Хищники и их обычные жертвы мирно жили вместе.
Другой пример условно-рефлекторного преобразования инстинктивного поведения показал эксперимент с рыбками цихлидами, которым во время их первого нереста заменили икру на икру чужого вида. Когда вывелись мальки, рыбки стали о них заботиться и оберегать, а когда в следующий нерест они вывели мальков своего вида, то гнали их как чужих. Таким образом, выработанные условные рефлексы оказались весьма консервативными. На основе подкрепления пищевыми и оборонительными реакциями у рыб вырабатывали различные двигательные условные рефлексы. Например, золотую рыбку учили проплывать через кольцо, делать «мертвые петли», бойцовая рыбка бетта блестящая, приученная проходить через отверстие в преграде, стала прыгать в него даже когда оно было поднято над водой.
Поведение рыб, их безусловные и условные рефлексы в значительной мере определяются экологическими факторами среды обитания, которая накладывает свой отпечаток на развитие нервной системы и формирования ее свойств.
Выработка оборонительных условных рефлексов у мальков. Регулирование течения рек, строительство плотин гидроэлектростанций и мелиоративных систем в большей или меньшей степени затрудняет путь рыбы к естественным нерестилищам. Поэтому все большее хозяйственное значение приобретает искусственное рыборазведение.
Ежегодно миллиарды мальков, выведенных на рыбоводных станциях, выпускаются в озера, реки и моря. Но только небольшая часть их выживает до промыслового возраста. Выращенные в искусственных условиях, они нередко оказываются плохо приспособленными к жизни на воле. В частности, мальки, не имевшие жизненного опыта образования защитных реакций, легко становятся добычей хищных рыб, от которых они и не пытаются спасаться. Для того чтобы повысить выживаемость мальков, выпускаемых рыбоводными станциями, были предприняты опыты по искусственной выработке у них защитных условных рефлексов на приближение хищных рыб.
В предварительных пробах изучали свойства образования таких рефлексов на зрительные, слуховые и вибрационные сигналы. Если среди мальков плотвы поместить металлические блестящие пластинки, имеющие форму тела хищника-щуренка, и пропускать через эти пластинки ток, то мальки начинают избегать этих фигур даже в отсутствие тока. Рефлекс вырабатывается очень быстро (рис. 84).
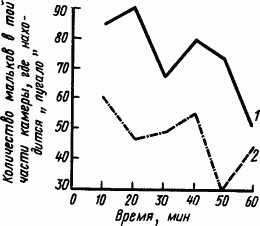
Рис. 84. Выработка условного оборонительного рефлекса у мальков плотвы на вид модели хищной рыбы в течение 1 ч (по Г.В. Попову):
1 — 35-дневные мальки, 2 — 55-дневные
Чтобы оценить, насколько выработка искусственных оборонительных рефлексов может повысить выживаемость молоди, сравнивали скорость поедания хищником мальков, прошедших подготовку, и мальков, не имевших такой подготовки.
Для этого в пруду устанавливались садки. В каждый садок помещали одну хищную рыбу — голавля и точно отсчитанное количество мальков рыбца. Через 1 или 2 сут подсчитывали, сколько мальков осталось в живых и сколько было съедено хищником. Оказалось, что из числа мальков, не вырабатывавших оборонительные рефлексы, почти половина гибнет в течение первых суток. Примечательно, что вторые сутки практически мало что добавляют в этом отношении. Можно думать, что уцелевшие мальки успевают образовать натуральные условные оборонительные рефлексы и успешно спасаются от преследований хищника. Действительно, если их брать после такой естественной подготовки в специальные опыты, то процент гибели оказывается или сравнительно небольшим, или даже нулевым.
Мальки с искусственно выработанными условными оборонительными рефлексами как на вид фигуры хищной рыбы, так и на сотрясение воды, имитирующее ее движения, меньше всего пострадали от голавля. В большинстве опытов хищник даже в течение двух суток не сумел изловить ни одного из них.
Разработанная в последнее время простая техника воспитания защитных рефлексов у мальков промысловых рыб во время их выращивания может принести существенную практическую пользу делу рыборазведения.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
§ 25. Теории происхождения хордовых
§ 25. Теории происхождения хордовых Бытует несколько точек зрения на возникновение хордовых, которые различаются как по подходам к решению проблемы, так и по животным, избранным в качестве представителя предковых групп. Наиболее известны гипотезы возникновения хордовых
§ 26. Происхождение нервной системы хордовых
§ 26. Происхождение нервной системы хордовых Наиболее часто обсуждаемые гипотезы происхождения не могут объяснить появление одного из основных признаков хордовых — трубчатой нервной системы, которая располагается на спинной стороне тела. Мне хотелось бы использовать
Глава 10 Нервная система
Глава 10 Нервная система ГипнотизмДругая разновидность заболеваний, которые не подпадают под теорию Пастера, — это заболевания нервной системы. Такие заболевания смущали и пугали человечество испокон веков. Гиппократ подходил к ним рационалистично, однако большинство
Глава 7 АНАЛИТИКО-СИНТЕТИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ МОЗГА
Глава 7 АНАЛИТИКО-СИНТЕТИЧЕСКАЯ ДЕЯТЕЛЬНОСТЬ МОЗГА Вся высшая нервная деятельность состоит из непрерывного анализа — расчленения раздражителей окружающего мира на все более простые их элементы, и синтеза — обратного слияния этих элементов в целостное восприятие
Глава 13 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЗЕМНОВОДНЫХ, ПРЕСМЫКАЮЩИХСЯ И ПТИЦ
Глава 13 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЗЕМНОВОДНЫХ, ПРЕСМЫКАЮЩИХСЯ И ПТИЦ Современные потомки первых обитателей суши сохранили в своей организации и поведении многие следы той ломки, которая сопровождала выход животных из водной стихии. Это видно, например, при
Глава 14 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ГРЫЗУНОВ И КОПЫТНЫХ
Глава 14 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ГРЫЗУНОВ И КОПЫТНЫХ После катастрофического конца эры холоднокровных гигантов, не сумевших приспособиться к новым условиям жизни, господствующее положение в животном мире заняли теплокровные млекопитающие. Высокий уровень обменных
Глава 15 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ХИЩНЫХ
Глава 15 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ХИЩНЫХ В жизни хищников приспособительное значение высшей нервной деятельности проявляется особенно ярко в жестокой борьбе за существование. Помимо непрерывной выработки все новых условных рефлексов защиты от более сильных врагов,
Глава 16 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ОБЕЗЬЯН
Глава 16 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ОБЕЗЬЯН Изучение высшей нервной деятельности обезьян представляет особый интерес по двум причинам. По-первых, обезьяны — самые высокоразвитые в психическом отношении животные, во-вторых, они самые близкие к человеку представители
Глава 17 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА
Глава 17 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА Жизнь на каждом шагу показывает неизмеримое превосходство разума людей над примитивными мыслительными способностями животных. Громадный разрыв между психической жизнью человека и животных долгое время служил поводом для
Глава XIII Нервная система
Глава XIII Нервная система Функции У нервной системы живых существ имеются две основные функции. Первая — сенсорное восприятие, благодаря которому мы воспринимаем и постигаем окружающий мир. По центростремительным чувствительным нервам импульсы от всех пяти органов
Эволюционные изменения в организации яиц хордовых
Эволюционные изменения в организации яиц хордовых Для Spiralia характерно развитие, при котором отдельные бластомеры на ранних стадиях дробления уже запрограммированы к дифференцировке в определенном направлении, не изменяющемся даже в том случае, если изолировать их от
Высшая нервная деятельность
Высшая нервная деятельность За 20–25 дней до начала опытов была сделана попытка охарактеризовать основные особенности нервных процессов каждой подопытной собаки, для чего проводились обследования с помощью проб, подробно описанных на с. 90 этой книги. В силу