Глава 16 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ОБЕЗЬЯН
Глава 16
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ОБЕЗЬЯН
Изучение высшей нервной деятельности обезьян представляет особый интерес по двум причинам. По-первых, обезьяны — самые высокоразвитые в психическом отношении животные, во-вторых, они самые близкие к человеку представители современного животного мира. Мозг обезьяны — это конечный результат эволюции биологических приспособлений и вместе с тем исходный материал, из которого труд мог создать орган человеческой мысли.
Прогресс обезьян определился прежде всего развитием у них рук. Рука дала им возможность хватать и подвергать тщательному исследованию все встречающиеся предметы. Отсюда «любопытство» обезьян, немедленно начинающих вертеть в руках все, что им попадается на глаза.
Условные рефлексы обезьян достигли высокого совершенства, особенно в отношении синтеза сигналов, полноты и целостности восприятия окружающего. Поэтому обезьяны далеко опередили всех других животных по своей «впечатлительности». Когда медвежонка, собаку и обезьяну привели в кинозал, где демонстрировался фильм «Хищные», то медвежонок, очутившись в темноте, закрыл глаза и задремал, собака больше оглядывалась назад на шум киноаппарата, а обезьяна не отрывала глаз от экрана. При появлении на экране тигра она схватилась за голову и с отчаянным воплем бросилась бежать.
Рефлексы детеныша обезьяны
Новорожденная обезьяна умеет лишь сосать, отстраняться от источника вредящих раздражений и цепляться за мать всеми четырьмя конечностями. Особенно резко выражены и сильно развиты у нее рефлексы цепляния, отражающие образ жизни лазающих животных.
Рефлексы цепляния. Рефлексы цепляния проявляются раньше других. Вышедший до половины из родовых путей детеныш макаки уже в состоянии крепко ухватиться за шерсть живота матери и помочь своему окончательному выходу на свет. Сразу после рождения детеныш так крепко цепляется за мать, что она может совершать свои обычные прыжки по деревьям. Если новорожденную обезьяну положить на гладкую поверхность, на которой не за что зацепиться, она кричит, рвется и размахивает руками. Как только ей удается во что-нибудь вцепиться — замолкает и успокаивается.
С возрастом цепляние переходит в схватывание. Схватывание быстро связывается с пищевыми подкреплениями и дает начало разнообразным натуральным условным рефлексам добывания пищи. Такие рефлексы также могут образовываться уже в первые дни жизни. Например, достаточно 10-дневному детенышу обезьяны однажды пососать кулачок, в котором оказалась случайно схваченная со стола соска, чтобы он, увидев соску, стал ее хватать и тянуть в рот. В последующие дни можно было наблюдать генерализацию этого новообразованного пищевого рефлекса — детеныш тащил в рот все, что мог схватить. Затем наступала специализация рефлекса, и он стал дифференцировать другие предметы от соски.
Сосательные, мигательные, пищевые и оборонительные рефлексы. Своеобразные условные сосательные рефлексы образуются у детенышей обезьян при искусственном вскармливаниии. Когда приближается время очередного кормления, они начинают что-нибудь сосать. Чаще всего это бывает какой-нибудь определенный палец собственной конечности.
У 10-дневного детеныша макаки можно выработать условный мигательный рефлекс на стук метронома, сопровождая его струей воздуха, направляемой в глаз.
В возрасте трех недель детеныш уже проявляет много натуральных пищевых и оборонительных рефлексов, хорошо дифференцирует приближающихся к нему людей, идет на руки к ухаживающим за ним и спасается бегством от посторонних.
Выработка суточного ритма активности. В 2–3-недельном возрасте у детенышей обезьян вырабатываются своеобразные условные рефлексы на время, из которых складывается ритм суточной активности. У новорожденного детеныша двигательная активность равномерно распределяется на протяжении суток. Лишь по мере выработки разнообразных пищевых и других условных рефлексов, приуроченных к дневному времени, складывается чередование активности в дневные часы и сна в ночное время.
Чесательные рефлексы. Еще на 3–4-м дне жизни у детенышей обезьян появляется чесательный рефлекс, являющийся основой выработки характерной для обезьян реакции «обыскивания». Такую реакцию у детенышей в возрасте 4–6 месяцев вырабатывают взрослые обезьяны. Они хватают детеныша, распластывают на полу и начинают обыскивать его шерсть — пальцами перебирают волоски, скребут кожу, выбирают грязь, перхоть, насекомых. Постепенно обыскивание приобретает у детенышей характер условной реакции по зрительным сигналам. Стоит одному из них подойти и подставить типичным жестом какую-нибудь часть тела, как другой немедленно начинает его обыскивать.
Память. Детеныши обезьян с раннего возраста могут запоминать значение отдельных сигналов в природе и в стаде путем образования на них условных рефлексов. Лишь постепенно развивается способность помнить картины событий в целом.
Развитие анализаторов обезьян
В окружающей обстановке обезьяны ориентируются главным образом зрительными и мышечными сигналами. Обоняние имеет ведущее значение. Этому способствовали питание легко доступными плодами и условия жизни на деревьях, куда плохо доносятся запахи следов с земли и откуда хорошо видно вокруг. Точный глазомер наряду с безошибочной двигательной координацией развился благодаря повседневной практике передвижения прыжками между ветвями.
Зрительный анализатор. Обезьян, как и птиц, называют животными «зрительного типа» в отличие от «обонятельного типа» хищников и «слухового типа» грызунов. Известно, что обезьяна плохо откликается на свою кличку и гораздо лучше на «пригласительный жест». Ее легче чему-нибудь выучить, используя в качестве сигналов главным образом мимику и жесты, а не приказы голосом.
Обезьяна обладает чрезвычайно острым зрением. Она может разглядеть черную нитку диаметром 0,2 мм на расстоянии 0,25 м даже при очень слабом освещении. Если поставить перед обезьяной пять прямоугольных ящиков, немного отличающихся друг от друга по вышине, и в один из них класть корм, то обезьяна научится различать его даже при разнице высоты в 25 мм. Обезьяны различали прямоугольники, площадки которых относились друг к другу как 1,16:1. Предел различения наступил, когда это отношение достигло 1,06:1.
Обезьяны анализируют детали зрительной картины лучше всех других животных. Очень тонкий анализ размеров и форм рассматриваемых предметов обнаружили шимпанзе в опытах с так называемым «выбором на образец» (Н. Ладыгина-Котс). В этих опытах экспериментатор показывал обезьяне один из лежащих перед ней предметов и добивался, чтобы обезьяна нашла его среди остальных и подала ему. В таком случае образуется цепь условных рефлексов на вид этого предмета, который связывается с двигательной реакцией схватывания. Последняя, в свою очередь, подкрепляется ориентировочно-исследовательскими и пищевыми реакциями. Оказалось, что шимпанзе отличала круг от овала, квадрата, пяти-, восьми-, десяти- и даже двенадцатиугольника. Бинокулярное зрение позволяет обезьянам оценивать перспективу и объемные формы. В этих же опытах шимпанзе прекрасно различали такие стереометрические фигуры, как шар, куб, призма, конус. Они отличали конус от трех-, четырех- и шестигранной призмы.
У обезьян, по-видимому, имеется настоящее цветовое зрение. Они могут, очевидно, различать длины световых волн, а не только яркость света. Убедительные доказательства цветового зрения у обезьян получены в опытах с мартышками, которые должны были находить пищу в стакане по цвету покрывавшей его бумаги.
Стаканы меняли местами и плотно закрывали, чтобы ни расположение стаканов, ни запах не могли указать, какой из них содержит пищу. Сигнальное значение яркости исключали заменой цветных бумаг серыми. Обезьяны хорошо различали двух- и трехцветные сочетания, т.е. легко образовывали из цветов комплексные условные раздражители.
Слуховой и обонятельный анализаторы. Обезьяны сравнительно мало пользуются слухом. Если не подкреплять словесную команду энергичными жестами, то обезьяну трудно заставить ее выполнить. Обратное развитие претерпело и обоняние обезьяны. Ее способность различать запахи не идет ни в какое сравнение с чутьем, например, собаки. Чтобы обнюхать предмет, обезьяна подносит его к морде.
Двигательный анализатор. Особое место в восприятии обезьяны занимают сигналы двигательного анализатора. Их роль в различении предметов по сравнению со зрительными и температурными сигналами показывают следующие опыты с шимпанзе.
В ящик на глазах у обезьяны запирали фрукты, рядом клали две металлические палки, одна из которых играла роль ключа. Эта палка была синего цвета, теплая и тяжелая. Другая палка была красного цвета, холодная и более легкая. После того как обезьяна научилась безошибочно выбирать нужную палку, ей предложили палки одинакового цвета и температуры, отличавшиеся только по массе. Обезьяна продолжала выбирать без ошибок палку, открывающую ящик. Когда же ей предложили палки одинаковой массы и температуры, отличавшиеся только по цвету, она начала ошибаться. Наконец, когда ей предложили палки одинаковой массы и цвета, различные только по температуре, обезьяна сделала еще больше ошибок. Следовательно, ведущими сигналами для дифференцирования у шимпанзе оказались кинестетические, затем шли зрительные, а за ними температурные.
Значение мышечных сигналов в жизни обезьян огромно. Обезьяна постоянно находится в движении. Когда она хватает, вертит и ощупывает все, что попадается в руки, то мышечные сигналы дают ей такие сведения о свойствах предмета, которые не получает никакое другое животное. На особое значение этих сигналов указывает также чрезвычайное развитие центрального отдела именно двигательного анализатора.
Условные слюноотделительные рефлексы. Описаны попытки применить для исследования высшей нервной деятельности обезьян классический метод условных слюнных рефлексов. У макак после 3–8 сочетаний звукового сигнала и пищевого подкрепления (конфеты) наблюдали условно-рефлекторное выделение слюны из околоушечной железы. Однако такое условное слюноотделение оказывалось очень нестойким и небольшим по величине (почти в 100 раз меньше безусловного). Натуральные условные слюнные рефлексы обезьян на вид пищи носят отпечаток их естественного способа питания плодами, срываемыми с деревьев.
Так, если приманку показывали издалека, то обезьяна лишь следила за ней взглядом, слюноотделение отсутствовало. Приближение приманки даже до 1 м и менее оставалось безрезультатным. Только когда приманка оказалась на расстоянии около 0,5 м, т.е. на расстоянии вытянутой руки, слюнные железы начинали работать.
Другая особенность слюнных натуральных условных рефлексов обезьян связана с тем, что в комплекс условного пищевого раздражения входят и сигналы от рук, которыми обезьяна хватает и обследует пищу. Результаты такого обследования в какой-то мере определяют и отношение к поедаемой пище. Поэтому, когда обезьяна не только видит, но и может ощупывать приманку, слюноотделение у нее резко возрастает. В одном из опытов у павиана анубис за 1 мин рассматривания лежащего перед ним плода выделилось 22 мл слюны, а за 1 мин рассматривания и ощупывания — 53 мл, т.е. слюноотделение увеличилось более чем вдвое.
Из-за беспокойного поведения обезьян с ними трудно работать по слюноотделительной методике, поэтому свойства их внешней нервной деятельности изучали главным образом по проявлениям различных двигательных условных рефлексов.
Двигательные оборонительные рефлексы. Простые двигательные оборонительные рефлексы отдергивания ноги при электрокожном раздражении изучали на зеленых мартышках. Во время опыта обезьян фиксировали в специальном станке с приспособлениями для регистрации дыхания и движения ноги.
Сочетания светового сигнала (красный цвет) с безусловной оборонительной реакцией (10 с — изолированное действие сигнала и 10 с — совпадающее) после 4–6 повторений приводили к выработке условного дыхательного рефлекса в виде резкого усиления дыхания. Однако условный рефлекс отдергивания ноги не образовался даже после 116–185 сочетаний.
Трудность выработки условного рефлекса отдергивания ноги, по-видимому, зависела от того, что очень подвижному животному, каким является обезьяна, более свойственна защитная реакция убегания. Лишенная возможности бегства, фиксированная в станке обезьяна реагировала на боль защитной неподвижностью.
Когда обезьяны имели возможность спасаться от болевого раздражения бегством, то двигательные оборонительные рефлексы вырабатывались легко. Так, условное бегство на верхнюю полку клетки по сигналу, вслед за которым нижняя часть клетки окатывалась струей воды, выработалось после 3–8 сочетаний. Условный рефлекс быстро упрочивался и был стойким. Без всяких затруднений можно было выработать к нему условный тормоз и образовать на его основе условный рефлекс второго порядка.
Двигательные пищевые рефлексы. У низших обезьян вырабатывали условные пищевые рефлексы подбегания к кормушке. По сигналу обезьяна (гамадрил) получала пищевое подкрепление в кормушке другой камеры, куда она должна была перебегать через качающийся люк. Первый условный рефлекс на метроном с частотой 104 удара/мин образовался после 11 сочетаний, второй условный рефлекс на тон — после 6 сочетаний. Дифференцировка метронома с частотой 60 ударов/мин выработалась после 14 применений. Однако рефлексы сильно тормозились посторонними случайными раздражителями. Ярко было выражено также последовательное торможение от дифференцировочного сигнала метронома.
Подобные опыты проводили также с резусами и с высшими обезьянами — орангутаном и шимпанзе. В этих опытах обнаружились сравнительно более быстрая выработка условных рефлексов и более короткое последовательное торможение у антропоидов. Можно также заключить, что у высших обезьян лучше идет концентрирование нервных процессов. У орангутана удалось наблюдать положительную индукцию при интервалах 30–45 с.
В других опытах у высших обезьян (шипманзе) вырабатывали условные пищевые реакции нажатия на клавиши своеобразного «обезьяньего рояля».
Определенная клавиша включала звонок, и тогда открывалась кормушка с фруктами. Была легко достигнута дифференцировка звонков по тону. В ходе выработки условных рефлексов можно было отчетливо видеть постепенное уточнение условной реакции и устранение лишних движений. Если при первых пробах обезьяна беспорядочно бьет всей пятерней по клавише, иногда даже садится на нее, протягивая руки к кормушке, то в дальнейшем она аккуратно нажимает клавишу одним пальцем, а свободной рукой берет персик из открывшейся кормушки.
Обращает на себя внимание бурная двигательная реакция на первые неподкрепления условного рефлекса при его угашении или специализации. Когда за нажатием клавиши не последовало открытия кормушки, шимпанзе стала бить «рояль» кулаками, яростно колотила себя в грудь, бросалась на окружающих и, наконец, схватившись за голову, с криком покатилась по полу клетки.
Свойства условных рефлексов обезьян исследовали также методикой выработки хватательных условных движений, являющихся для этих животных естественными пищедобывательными актами. Подготовка животного к таким опытам заключалось в том, что вначале каждое случайное схватывание рычага подкрепляли подачей лакомого корма. Когда обезьяна прочно закрепляла это движение как средство добывания пищи, приступали к выработке разнообразных искусственных условных рефлексов на звуковые, световые и прочие сигналы. Для этого обезьяна получала пищевые подкрепления только тогда, когда она схватывала рычаг по сигналу. Межсигнальные схватывания оставались без подкрепления и вскоре угасали. Когда условный рефлекс образовался, то достаточно одного сигнала, чтобы обезьяна схватила рычаг, дернула за него и взяла из кормушки кусочек пищи, спущенный туда через желоб экспериментатором. Обычно каждый сигнал действовал около 10 с, интервалы между ними были нерегулярные — 1–5 мин, чтобы не образовался рефлекс на время.
Низшие обезьяны (макаки, мартышки, павианы) образовывали положительные условные рефлексы на слуховые и зрительные сигналы в пределах 2–8 сочетаний. Действуя, как гаснущие тормоза, случайные раздражения могут уменьшать и даже полностью задерживать выработанные условные рефлексы, создавая впечатление их непрочности и нерегулярности. В действительности условные рефлексы обезьян очень прочны. При отсутствии посторонних раздражителей они повторяются из опыта в опыт с большим постоянством и сохраняются при отсутствии практики гораздо дольше, чем у других животных.
Например, у одной макаки после длительного перерыва в работе при первом же испытании появились условные рефлексы на сложной стереотип раздражителей, выработанный 8 лет назад.
Низшие обезьяны весьма тонко дифференцируют зрительные и слуховые сигналы. Выработка дифференцировочного торможения достигалась после 10 применений подкрепляемых и неподкрепляемых раздражителей. Для выработки условного тормоза требовалось 8–10 применений комбинации из условного раздражителя и прибавочного агента. Угасание условного рефлекса происходило после 2–10 неподкреплений положительного условного раздражителя, повторенных подряд с интервалом 0,5–1 мин. Здесь также может сильно сказываться влияние посторонних раздражений, легко вызывающих растормаживание и создающих ложное впечатление слабости внутреннего торможения.
Условные рефлексы на сложные раздражители. Образование условных рефлексов на комплексные сигналы — цепи раздражителей и одновременные комплексы — происходит у обезьян легко, иногда даже быстрее, чем на одиночные. Так, на трехчленную цепь раздражителей (метроном + шум + свет лампочки) условный рефлекс образовался через 2–3 сочетания. В отличие от собак обезьяны сумели достичь абсолютной дифференцировки трехчленных цепей раздражителей по среднему члену цепи. У них раньше и лучше вырабатывалось запаздывание, приурочивающее условную реакцию к концу действия комплексного раздражителя. На одновременный комплекс раздражителей (звонок+метроном+белый свет или тон+шум+зеленый свет) условный рефлекс образовался с первых же сочетаний. Дифференцировка вырабатывалась после первых применений неподкрепляемого варианта комплекса. Наконец, чрезвычайно важной является способность обезьян к выработке условных рефлексов на следы давно бывших раздражений. Благодаря этой способности у них могут образоваться длинные цепи условных связей, и ряд вытекающих одно из другого действий складывается в картины сложного поведения. Во всем этом проявляется более высокий, чем у других животных, уровень аналитико-синтетической деятельности мозга обезьян.
Мозг обезьян может производить очень сложную аналитико-синтетическую работу, воспринимая комплексный раздражитель и его отдельные компоненты как сигналы для разных реакций.
Так, у павиана гамадрила был выработан условный пищевой рефлекс потягивания за рычаг на последовательный комплекс раздражителей (свисток + квадратная световая фигура). После укрепления пищевого рефлекса на комплекс в целом второй компонент комплекса (квадратная световая фигура) стали сочетать с оборонительной реакцией убегания на верхнюю полку клетки от струи воды, заливающей нижнюю часть клетки. Очень скоро, после 3–6 сочетаний, произошло полное переключение этого компонента на оборонительную реакцию (рис. 98, А). Таким образом, один и тот же раздражитель в составе комплекса воспринимался как часть сигнала пищи, а примененный самостоятельно — как сигнал опасности.
Другой тонкий аналитико-синтетический акт, недоступный собакам, связан с переключением условно-тормозного сигнала. Если сделать из стука метронома условный тормоз для ряда условных пищевых раздражителей (вертушка, свет лампочки и др.), а затем переключить свет лампочки с пищевой на оборонительную реакцию, то тормозное значение метронома сохранится для вертушки, но потеряется для лампочки (рис. 98, Б). Здесь ярко выступает целостное восприятие обезьяной раздражителя в связи со всеми обстоятельствами его действия.
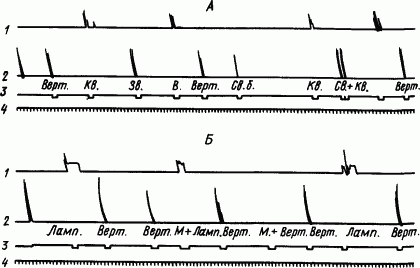
Рис. 98. Переключение сигнального значения условных раздражителей у обезьяны. А — полное расчленение сигнального значения комплекса и отдельно примененного его компонента; Б — устранение условно-тормозного значения агента после переключения сигнала на другую реакцию (по О.В. Малиновскому):
1 — запись оборонительной реакции вскакивания на верхнюю полку, 2 — запись пищевой реакции дергания за рычаг, 3 — отметки условных раздражений, 4 — отметка времени (5 с); сигналы пищевой реакции: Верт. — вертушка, Зв. — звонок, Св.б. — белый свет, Св. + Кв.— комплекс свисток+квадратная световая фигура, сигналы оборонительной реакции: Кв.— второй компонент комплекса, Ламп. — свет лампочки, М. — стук метронома
Способность обезьян к сложной аналитико-синтетической деятельности, обеспечивающей приспособление к новым условиям жизни, наглядно была продемонстрирована в наблюдениях за поведением группы макак-резусов, привезенных из Индии и выпущенных на лесистом острове Псковской области. Обезьяны настолько адаптировались к новым для них условиям климата, флоры и фауны, что сами обеспечивали себе питание, добывая естественную пищу, а процессы обучения происходили у них быстрее и эффективнее, чем в лабораторных условиях.
Сложное поведение обезьян формируется с раннего возраста и в значительной мере зависит от условий их жизни в стаде. С этим связано и их эмоциональное поведение, в том числе сексуальное, определяющее взаимоотношения особей в разных ситуациях.
Свойства условных рефлексов антропоидов
Наиболее высокого совершенства достигает аналитико-синтетическая деятельность мозга у высших человекообразных обезьян. Это подтверждают опыты с шимпанзе, которые в течение многих лет проводят в «столице условных рефлексов» — Колтушах (ныне с. Павлово) сотрудники Института им. И.П. Павлова. Об основных свойствах процессов высшей нервной деятельности антропоидов дают представление исследования хватательных условных рефлексов, вырабатываемых на пищевом подкреплении.
Первые условные положительные рефлексы на звуковые и световые сигналы появились у молодых шимпанзе после 5–8 сочетаний, скорость дифференцирования сигналов была больше подвержена колебаниям. Когда приступили к обратной переделке сигналов значения условных раздражителей, то она произошла быстрее, чем выработка их первоначального значения. Возврат к прежнему значению сигналов осуществился еще легче. Вскоре достаточно было только один раз подкрепить неподкрепляемый сигнал и не подкрепить подкрепляемый, чтобы они сразу поменяли свое значение на противоположное. В то время как для большинства животных многократная полная обратная переделка сигнального значения раздражителей является трудной задачей, для антропоидов эта операция не составляет никакого труда.
Эмоции антропоидов. В опытах с человекообразными обезьянами обращают на себя внимание своеобразные эмоциональные компоненты реакций на отрицательные сигналы. Зачатки таких эмоций наблюдались уже у низших обезьян. Например, павиан сфинкс при подаче отрицательного условного раздражителя кричит, делает «жесты угрозы». Выше описана бурная реакция «гнева» взрослого шимпанзе в ответ на дифференцировочный сигнал. Молодая, обычно очень спокойная шимпанзе при подаче дифференцировочного раздражения била кулаком по стеклу, за которым зажигалась сигнальная лампочка, кричала, отворачивалась. Особое «негодование» шимпанзе вызывала условно-тормозная комбинация сигналов. Обезьяна вначале застывала на несколько секунд в оцепенении, затем с гримасами и выразительными жестами бросалась к сигнальной доске, била ее кулаками, кидалась на пол. Повторение отрицательных сигналов усиливало бурную реакцию «протеста», которая дополнялась «жалобными» звуками и судорожным расчесыванием кожи, как будто обезьяну охватил нервный зуд.
Такие эмоциональные ответы на отрицательные раздражители, свойственные обезьянам, по-видимому, зависят от растормаживания подкорковых реакций при торможении коры. Интересно, что и маленькие дети при исследовании у них условных рефлексов отмахиваются от дифференцировочных сигналов, дуют на них, убегают из кабины. Очевидно, лишь с возрастом кора мозга человека настолько полно контролирует работу подкорковых механизмов, что почти при любом своем состоянии не допускает преобладания их деятельности.
Не менее выразительны и эмоциональные проявления реакций антропоидов на положительные сигналы. Из них особый интерес представляют мимические движения «смеха». Человекообразные обезьяны — единственные животные, которые умеют «смеяться от радости». Другое сходство эмоциональных проявлений у антропоида и человека заключается в живой игре сосудов кожи. Шимпанзе бледнеют и краснеют при своих «переживаниях».
Исследовательские рефлексы. Молодые шимпанзе легко образуют временные связи сочетаемых индифферентных раздражителей (например, светового и звукового) — то, что психологи называют ассоциацией по времени. Основой этой связи становятся ориентировочно-исследовательские реакции, так сильно развитые у обезьян. Однако, если такая связь не получает безусловного подкрепления, она быстро затормаживается. Это зависит от развития внутреннего торможения и угасания вначале резко выраженных ориентировочных рефлексов на ассоциируемые раздражители.
Ориентировочно-исследовательский инстинкт играет выдающуюся роль в жизни человекообразных обезьян. Вообще ориентировочная реакция установки рецепторов в сторону источника нового раздражителя свойственна всем животным. Но обезьяна (с ее хватающей и ощупывающей рукой) дополнила ориентировочную реакцию активной формой исследовательской реакции. Она схватывает все, что увидит, ощупывает, вертит, ковыряет, пробует на прочность, разглядывает со всех сторон. Ее подвижные пальцы, действуя вместе с внимательным взглядом, быстро пробегают по поверхности схваченного предмета, проникают в отверстия, пытаются разъединять и соединять его части. Расширяя сферу исследования, обезьяна обнюхивает, пробует на вкус, примеряет предметы друг к другу, прикладывает их к разным частям своего тела.
Развитие исследовательских условных рефлексов, направляющих животное на более активное вмешательство в окружающие события, видимо, явилось важным фактором биологического прогресса приматов.
Голосовые и мимические рефлексы. Стадная жизнь обезьян способствовала развитию у них голосовой, жестикуляторной и мимической сигнализации. Например, павианы могут издавать около 25 различных звуков, имеющих определенное сигнальное значение. Если стадные звуки общения павианов анубисов, имеющие характер цокания и бормотания, подкреплять пищей, то образуется голосовой условный пищевой рефлекс. По сигналу павиан цокает или бормочет и протягивает руку за подкормом. Осциллографическая регистрация позволяет дать точную фонетическую характеристику голосовым сигналам разного биологического значения. Так, у коричневых капуцинов отчетливо разделяются звуки, сигнализирующие различные виды ориентировочно-исследовательских, оборонительных и пищевых реакций. Очень распространена среди обезьян и сигнализация телодвижениями. У низших обезьян преобладают поза и жест. Примерами могут служить «поза покорности» у макак или «угрожающие жесты» павианов. У человекообразных обезьян развивается выразительная мимика, тонко отображающая «настроение», т.е. биологическую направленность общей установки поведения животного.
Подражательные рефлексы. Высокое совершенство исследовательских реакций, обстановка стадной жизни и чрезвычайная подвижность нервных процессов — все это создало благоприятные условия для особенного развития у обезьян подражательных условных рефлексов. Обыкновение обезьян слепо повторять увиденное, «обезьянничать» — общеизвестно. В развитии способности к подражанию они приобрели большое физиологическое преимущество использования чужого жизненного опыта. Существует много рассказов об умении обезьян учиться тому, что происходит у них на глазах.
Опыты с выработкой у обезьян искусственных подражательных условных рефлексов выявили некоторые особенности их свойств.
Например, у павиана гамадрила вырабатывали обычный условный двигательный рефлекс открывания кормушки на сигнал — тон аккордеона. На расстоянии 1–2 м от него в другой клетке находились обезьяны-«зрители», наблюдавшие за всеми действиями «актера». Когда у обезьяны-«актера» выработался условный рефлекс, на его место посадили одного из «зрителей». При первой же пробе сигнала «зритель» подбежал к кормушке и открыл ее, хотя сам никогда не имел случая производить по сигналу подобное действие. Следовательно, наблюдая за поведением «актера», «зрители» переняли выработанный им условный рефлекс. Примечательно, что к несколько иному сигналу — тон октавой ниже — «зритель» оставался безразличным, в то время как «актер» еще продолжал подбегать к кормушке. Следовательно, условный рефлекс «зрителя» оказался сразу специализированным, в то время как у «актера» он еще проходил стадию генерализации.
Интересный случай подражательного поведения обезьян описал японский исследователь М. Каваи, наблюдавший за жизнью макак на Кошима. Одна из них, найдя на берегу реки в песке батат, уронила его в воду, он очистился от песчинок и обезьяна его съела. В дальнейшем она стала пользоваться этим приемом, который привлек внимание и других макак и через некоторое время из 59 обследованных обезьян 42 мыли бататы перед едой.
Натуральные подражательные условные рефлексы играют громадную роль в поведении обезьян. Поведение вожака быстро перенимается членами семьи. Детеныши с самого раннего возраста учатся сложным защитным, пищевым, исследовательским и другим реакциям у взрослых животных. Например, искусственно вскормленные в лаборатории детеныши обнаруживали мимические и жестикулярные реакции только в 7–9-месячном возрасте, а выросшие в стаде — уже в возрасте 3–4 месяцев.
Ниже показана трудность выработки рефлекса у 4 обезьян гамадрилов подкатывания лопаткой персиков, чтобы их можно было достать рукой. Затем одна из них демонстрировала эту новую пищедобывательную реакцию четырем другим обезьянам в течение 3 дней по 5–6 раз в день. Когда этих обезьян тоже стали обучать подкатыванию персиков лопаткой, они научились это делать в пять раз быстрее, чем обезьяны, не проходившие такой предварительный подготовки.
Число проб, потребовавшихся для выработки рефлекса у обезьян, ранее не видавших такого движения / Число проб, потребовавшихся для выработки рефлекса у обезьян, присутствовавших при демонстрации рефлекса
52 /10
80 / 15
120 / 25
205 / 31
Нетрудно представить, насколько подражание облегчает обезьянам приспособление к разнообразной жизненной обстановке. Особой сложности достигают подражательные условные рефлексы у человекообразных обезьян. Развитие этих рефлексов, вероятно, сыграло немаловажную роль в их выдвижении на исключительное положение в животном мире.
Физиологический анализ сложного поведения человекообразных обезьян
Биологические предшественники человека — антропоиды — выделяются своим поведением среди всех других животных. Активная исследовательская деятельность, расширение жизненного опыта путем подражания, высокое развитие синтеза и целостное восприятие окружающего, наконец, чрезвычайно легкая обучаемость с использованием следов давних раздражений — все эти особенности их высшей нервной деятельности имеют большое сходство с некоторыми общими свойствами высшей нервной деятельности человека. Однако несмотря на сходство их психической, так же как и анатомической, организации даже самую высокоорганизованную обезьяну нельзя отождествлять с человеком.
Поведение человекообразных обезьян. Несмотря на то что некоторые действия антропоидов могут походить на человеческие поступки, к ним, как и вообще к действиям животных, даже способных к сложному поведению, называемому рассудочным, совершенно не применимы объяснения и понятия, заимствованные из психологии.
Особенности ума человека, его способность к отвлеченному мышлению возникли в результате общественного образа жизни, в корне отличающегося от биологического образа жизни даже высших животных. Поэтому нет оснований искать у последних сознательные идеи и разумные рассуждения.
Однако ошибочное перенесение на антропоида свойств человеческого сознания до сих пор еще довольно распространено. Оно основано, с одной стороны, на ярких впечатлениях от «умного» поведения обезьян, а с другой — на метафизическом игнорировании социально обусловленных особенностей сознания человека.
Действительно, высшие обезьяны, особенно при содержании в домашней обстановке, могут воспроизводить многие поступки человека.
Например, описывают, что молодой самец гориллы, живший в доме одного англичанина, входя вечером в комнату, включал электрический, свет, ел за столом, умел пользоваться кранами в ванной, спал в постели и укрывался одеялом.
В 1933 г. проф. С. Воронов прислал из Франции в подарок И.П. Павлову двух шимпанзе — брата и сестру, названных Рафаэлем и Розой. Для них в Колтушах выстроили специальное здание. И.П. Павлов с большим интересом наблюдал за поведением антропоидов. Он называл Рафаэля за пристрастие к еде «утробистым господином», а Розу за неутомимость исследовательских реакций — «интеллигентной особой». Шимпанзе быстро освоились со своей новой квартирой. Они обедали за столом, сидя на стульях, пользовались вилками, охотно принимали ванны.
Однако было нетрудно убедиться, что навыки «человеческого» поведения происходят не от потребностей обезьяны, а навязаны ей людьми. Когда заботливый воспитатель, уложив Рафаэля в кровать, уходил, то шимпанзе вскакивал, рвал матрац и из набивки устраивал себе гнездо на полу. Обедая за столом, с салфеткой, он норовил схватить кружку не рукой, а ногой. Не исключена также возможность, что любовь к ванной питалась не столько чистоплотностью, сколько пристрастием к мылу, которое Рафаэль охотно поедал.
Абстрактные понятия. Многие психологи экспериментально изучали сложное поведение высших обезьян, пытаясь определить, могут ли они мыслить абстрактными понятиями (идеями), как люди. Например, Р. Йеркс (1917), работая методикой «множественного выбора», сравнивал приемы решения задач низшими и высшими обезьянами.
Способна ли человекообразная обезьяна к абстрактным понятиям? В опытах, имеющих целью ответить на этот вопрос, обезьян выпускали из камеры в помещение, в конце которого были видны двери в 12 отделений. В каждом опыте была открыта часть этих дверей, в том числе одна, ведущая в отделение, где обезьяну ждала лакомая пища. Из опыта в опыт двери открывали в разных комбинациях, но дверь, ведущая к пище, занимала в этой комбинации всегда определенное место. Смысл исследования заключался в том, чтобы определить, сумеет ли обезьяна отвлечься от частностей конкретного расположения нужной двери в каждом отдельном опыте и овладеть общей идеей ее положения в комбинации открытых дверей. Например, в одном опыте были открыты двери 2, 3, 4, 5-я, из них вела к пище — 3-я, в другом опыте были открыты двери 8, 9, 10-я, к пище вела — 9-я, в третьем опыте открыты 4, 5, 6, 7, 8, 9-я, к пище вела — 5-я и т.д. Обезьяна должна была сделать обобщение — к пище ведет 2-я дверь слева из числа открытых.
Результаты опытов с мартышками и орангутаном представлены в виде «кривых научения» (рис. 99): эти кривые различны. У мартышек обучение происходит постепенно, как у всех животных, путем «проб и ошибок», т.е. вырабатывался все более специализированный условный двигательный рефлекс выбора заданной двери путем пищевого подкрепления правильных и неподкрепления не приводящих к результату движений. Иначе выглядит «кривая научения» орангутана. В течение длительного времени он не обнаруживал никаких успехов. Даже тогда, когда мартышки после 100–150 опытов уже решили задачу, он делал все те же ошибки. Более того, в дальнейшем число ошибок даже увеличилось. И только на 300-м опыте вдруг наступил крутой перелом — орангутан сразу решил задачу. Из внезапности решения задачи был сделан вывод, разделяемый многими психологами, что в отличие от мартышек он пришел к этому решению путем внезапной «догадки» об идее задачи.
Однако такое приписывание обезьяне свойств человеческого ума глубоко неверно, а полученные в опыте результаты имеют более естественное физиологическое объяснение. Это объяснение вытекает из описанных в предыдущей главе особенностей высшей нервной деятельности антропоидов.
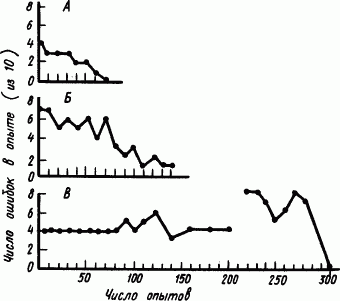
Рис. 99. Кривые «научения», полученные на мартышках (А, Б) и орангутане (В) в опытах с «множественным выбором» (по Р. Йерксу)
Конкретные рефлексы, а не абстрактные понятия. Как известно, человекообразные обезьяны отличаются необычайным развитием исследовательских реакций, которые могут даже подавлять пищевые. Недаром для работы с обезьяной в «проблемной клетке» побуждением чаще служит не пища, за которой она должна проникнуть в клетку, а возможность открыть сложный затвор, чтобы выйти из клетки. По-видимому, и в опытах со множественным выбором активная исследовательская деятельность орангутана долгое время задерживала выработку условного пищевого рефлекса правильного выбора двери, ведущей к пище. Когда же нужное движение связалось с пищевым подкреплением, то проявились свойственные антропоидам быстрота и прочность закрепления временных связей. Все это создало впечатление внезапного решения задачи.
В действительности обезьяны, как и все животные, руководствуются в своем поведении не абстрактными понятиями, идеями, а конкретными сигналами действующих на них условных раздражителей. Об этом убедительно свидетельствует следующий опыт.
Рафаэля обучили доставать фрукты из шкафчика, перед дверцей которого горел огонь. Для этого он должен был набирать из стоящего неподалеку бака воду в банку и забивать пламя, после чего мог взять и съесть приманку. В жаркие летние дни Рафаэля вывозили на озеро, и он проводил время на плоту. Когда солнце припекало очень сильно, он ложился на край плота, зачерпывал банкой из озера воду и обливался. Казалось бы, у Рафаэля должно сложиться общее понятие о воде и ее свойствах. Но вот на плоту Рафаэля установили шкафчик с огнем, препятствующим доступу к фруктам, а на соседний плот поставили бак с водой. Увидев знакомую ему аппаратуру, Рафаэль схватил банку, подбежал к краю плота, но вместо того, чтобы зачерпнуть из озера находящуюся перед ним воду, начал сооружать из лежащих рядом шестов мостик на соседний плот. С трудом перебравшись по этому шаткому мостику, он наполнил банку водой из бака и осторожно вернулся с ней на свой плот. Не один раз проделывал Рафаэль такое путешествие, пока ему удалось погасить огонь и завладеть фруктами.
Рафаэль не воспользовался для тушения огня уже знакомой ему водой из озера, потому что он не имел абстрактного понятия о воде и ее свойствах. В каждом частном случае вода выступала перед ним как конкретный раздражитель иного значения. Вода бака — компонент комплексного сигнала заливания огня в шкафчике с фруктами, т.е. условного пищедобывательного рефлекса. Вода озера — компонент комплексного сигнала охлаждения разгоряченного тела, т.е. условного теплорегуляторного рефлекса. В этом принципиальное отличие мышления антропоида от сознания человека, в котором складываются отвлеченные понятия, например идея о воде с ее общими свойствами, абстрагированная от частных проявлений этих свойств.
Процессы мышления антропоидов
Обширные исследования «интеллекта» человекообразных обезьян проводили на Биологической станции Прусской Академии наук на о. Тенериф (В. Келер, 1930). Методикой анализа решений различных сложных задач и действий в необычных положениях изучали процессы мышления шимпанзе. Результаты этих очень интересных опытов рассматривали в духе гештальт-психологии, о которой уже шла речь в связи с вопросом о системности работы мозга. По мнению гештальтистов, обезьяна, увидев за решеткой клетки виноград, достает его палкой не потому, что в ее мозгу уже имеется связь вида палки с пищевым подкреплением, образованная материальным действием внешних раздражителей, а потому, что в голове обезьяны вдруг возник соответствующий образ — зрительная идея. Неправомерная даже для понимания психики человека, гештальт-психология была приложена к человекообразным обезьянам. Фактическая необоснованность такого подхода к изучению психики антропоидов была показана в многочисленных контрольных экспериментах, проведенных в Институте физиологии им. И.П. Павлова с шимпанзе Рафаэлем и Розой (Э.Г. Вацуро, 1948).
«Ага-реакция». Одно из главных доказательств того, что шимпанзе решали задачи якобы в результате внезапного озарения их ума идеей, усматривалось в так называемой «ага-реакции». Например, обезьяна безуспешно пыталась достать высоко подвешенный плод. Она вытягивала то одну, то другую руку, прыгала на месте и с разгона, но схватить его не могла. Тогда она отошла в сторону и некоторое время сидела неподвижно, потом вдруг вскочила, побежала в угол клетки, где стоял ящик для игры, подтянула его под висящую приманку, взобралась на ящик и достала лакомство. Психологи-гештальтисты утверждают: убедившись в бесполезности действий наугад, обезьяна стала размышлять. Перед ее «умственным взором» мелькали «элементы зрительного поля», ее «ум» находился в состоянии «предгештальтного напряжения». Вдруг наступило «прояснение» образа, возник нужный гештальт, обезьяну «осенила идея» воспользоваться ящиком. Отсюда и название реакции — «ага, догадался!».
Физиологическое объяснение «ага-реакции». Для объяснения «ага-реакции» нет необходимости обращаться к понятиям, заимствованным из категорий сознания человека. Двигательные реакции, вызванные видом лакомой пищи, как и всякие неподкрепляемые условные рефлексы, угасали. Обезьяна успокаивалась, и ее поза создавала впечатление размышлений. Тогда начинали освобождаться от отрицательной индукции, созданной двигательным возбуждением, многие другие ранее образованные связи. И когда растормозилась связь вида ящика с реакцией влезания на него, уже образованная в неоднократных играх, шимпанзе бросилась к нему и осуществила эту реакцию. Что обезьяна действовала по выработанным связям, а не в результате «озарения идеей», ясно видно из того, что после безуспешных попыток достать плод, лежащий на земле за решеткой, она также бросилась к ящику, хотя это было совершенно бессмысленным поступком.
«Обходные пути». Вторым сильным доказательством наличия заранее обдуманных идей в действиях шимпанзе считали результаты опытов, которые обычно называют испытаниями на «обходы». Дело в том, что, как правило, животные, увидев пищу, стремятся к ней напрямик. Например, если перед клеткой с курицей рассыпать зерно, то она будет пытаться просунуть голову через решетку, биться об нее, но не воспользуется открытой дверцей в противоположной стенке. Если же на глазах у шимпанзе, находящегося в комнате, бросить за окно банан, то обезьяна немедленно побежит прочь от окна к двери. Она выскочит через коридор в выходную дверь наружу и устремится под окно, где начнет искать брошенный плод. Такое поведение обезьяны якобы указывает на то, что она в отличие от других животных держит в своей голове общее представление о плане дома и поэтому представляет себе, что путь к брошенному за окно банану лежит через удаление от окна и обходный путь по коридору.
Подобные факты наблюдали также в опытах с так называемым «обходным щитом». Это невысокая загородка, расположенная перед клеткой с шимпанзе и открытая в противоположную сторону. В загородку кладут банан, а обезьяне дают палку. После нескольких безуспешных попыток пододвинуть банан к клетке шимпанзе научается сначала толкать плод от себя, и только когда он выйдет за пределы загородки, поворачивать его по направлению к клетке. Такое решение задачи расценивается как возникновение у шимпанзе «идеи обхода».
Физиологическое объяснение «реакции обхода». Факты обучения обходным действием совсем не требуют для своего объяснения предположения о возникновении «идеи», а полностью укладываются в известные свойства цепных двигательных условных рефлексов. Более того, подобные реакции можно наблюдать и у других животных с менее развитой психикой. Например, дворовая собака, увидев через забор на улице кошку, бросается «в обход» к воротам. Следовательно, «реакция обхода» не является исключительным приобретением и свидетельством отвлеченного мышления антропоидов. Что касается результатов опытов с «обходным щитом», то они объясняются выработкой цепей двигательных пищедобывательных рефлексов. Недаром шимпанзе «осеняет идея» обхода лишь после того, как ему однажды удается достать плод, случайно оказавшийся поблизости от выхода из загородки, т.е. после пищевого подкрепления такого действия. Отсутствие при этом у шимпанзе сознательной «идеи обхода» убедительно показывает контрольный вариант этих опытов, проведенный в Колтушах с Рафаэлем.
После того как Рафаэль научился извлекать банан из загородки обходом (рис. 100), в загородке прорезали прямой выход в сторону клетки. Если бы обезьяна была способна к сознательной оценке обстановки, то «идея обхода» должна была бы сразу отпасть. Однако этого не произошло. Рафаэль продолжал бессмысленные теперь обходы вместо легкого движения к себе, которым он прямо мог получить банан в руки. Следовательно, обход не был осуществлением его отвлеченной идеи, а являлся конкретным пищедобывательным условным рефлексом, быстро образованным на пищевом подкреплении и прочно закрепленным, как это свойственно высшей нервной деятельности антропоидов.
О механическом действии этого рефлекса вопреки требованиям сознательного разума, свидетельствует факт движения в обход, даже когда банан лежит прямо на выходе из загородки к клетке (рис. 100).

Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 10 Нервная система
Глава 10 Нервная система ГипнотизмДругая разновидность заболеваний, которые не подпадают под теорию Пастера, — это заболевания нервной системы. Такие заболевания смущали и пугали человечество испокон веков. Гиппократ подходил к ним рационалистично, однако большинство
Глава 12 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЛИЧИНОЧНО-ХОРДОВЫХ КРУГЛОРОТЫХ И РЫБ
Глава 12 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЛИЧИНОЧНО-ХОРДОВЫХ КРУГЛОРОТЫХ И РЫБ Высшая нервная деятельность позвоночных животных отражает одну из важных тенденций их эволюции — индивидуальное совершенствование. Эта тенденция проявляется в нарастающей продолжительности
Глава 13 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЗЕМНОВОДНЫХ, ПРЕСМЫКАЮЩИХСЯ И ПТИЦ
Глава 13 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЗЕМНОВОДНЫХ, ПРЕСМЫКАЮЩИХСЯ И ПТИЦ Современные потомки первых обитателей суши сохранили в своей организации и поведении многие следы той ломки, которая сопровождала выход животных из водной стихии. Это видно, например, при
Глава 14 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ГРЫЗУНОВ И КОПЫТНЫХ
Глава 14 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ГРЫЗУНОВ И КОПЫТНЫХ После катастрофического конца эры холоднокровных гигантов, не сумевших приспособиться к новым условиям жизни, господствующее положение в животном мире заняли теплокровные млекопитающие. Высокий уровень обменных
Глава 15 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ХИЩНЫХ
Глава 15 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ХИЩНЫХ В жизни хищников приспособительное значение высшей нервной деятельности проявляется особенно ярко в жестокой борьбе за существование. Помимо непрерывной выработки все новых условных рефлексов защиты от более сильных врагов,
Глава 17 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА
Глава 17 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЧЕЛОВЕКА Жизнь на каждом шагу показывает неизмеримое превосходство разума людей над примитивными мыслительными способностями животных. Громадный разрыв между психической жизнью человека и животных долгое время служил поводом для
Глава XIII Нервная система
Глава XIII Нервная система Функции У нервной системы живых существ имеются две основные функции. Первая — сенсорное восприятие, благодаря которому мы воспринимаем и постигаем окружающий мир. По центростремительным чувствительным нервам импульсы от всех пяти органов
Высшая нервная деятельность
Высшая нервная деятельность За 20–25 дней до начала опытов была сделана попытка охарактеризовать основные особенности нервных процессов каждой подопытной собаки, для чего проводились обследования с помощью проб, подробно описанных на с. 90 этой книги. В силу
Предметная деятельность обезьян
Предметная деятельность обезьян Свое конкретное воплощение взаимодействие зрения и тактильно-кинестетической чувствительности рук находит в чрезвычайно интенсивной и многообразной манипуляционной активности обезьян.Исследования, проведенные рядом советских
Предтрудовая предметная деятельность ископаемых обезьян
Предтрудовая предметная деятельность ископаемых обезьян Не переоценивая орудийную деятельность современных антропоидов, нельзя одновременно не усмотреть в ней свидетельство одной из важных биологических предпосылок антропогенеза.Надо думать, что у ископаемых
6. Величины IQ обезьян, человекообразных обезьян и гоминид до Homo sapiens
6. Величины IQ обезьян, человекообразных обезьян и гоминид до Homo sapiens Делались попытки оценить интеллект обезьян, человекообразных обезьян и гоминид до Homo sapiens, основываясь на теории Пиаже о развитии интеллекта у детей. Согласно теории Пиаже, дети проходят четыре стадии
Глава 9. Обезьянолюди и другие странные существа Когда и почему люди отделились от обезьян?
Глава 9. Обезьянолюди и другие странные существа Когда и почему люди отделились от обезьян? Конечно, эволюция человека не ограничилась изменением волосяного покрова, молочных желез и плаценты. Мы в конце концов приматы, «первые» – пусть это и с трудом можно было
Глава 13. На заре любви: половое поведение обезьян
Глава 13. На заре любви: половое поведение обезьян Социальные отношения у обезьян: различия между полами Гипотеза пресса хищников и гипотеза межгрупповой конкуренции за пищевые ресурсы сходятся в одном важном пункте: они признают, что виды существенно различаются, в