Эволюционные изменения в организации яиц хордовых
Эволюционные изменения в организации яиц хордовых
Для Spiralia характерно развитие, при котором отдельные бластомеры на ранних стадиях дробления уже запрограммированы к дифференцировке в определенном направлении, не изменяющемся даже в том случае, если изолировать их от остального зародыша. Широкое распространение получили представления о том, что такое мозаичное развитие типично для первичноротых, тогда как в основе развития вторичноротых лежат индукционные взаимодействия между клетками, детерминирующие их судьбу. Эти представления неверны по двум причинам. Во-первых, у некоторых вторичноротых, в особенности у асцидий, развитие столь же высокомозаичное, как и у любого представителя Spiralia. Во-вторых, детерминированность определенных клеток к развитию по определенному пути содержит в себе временной фактор. У тех зародышей, которых принято считать типично мозаичными, эта детерминированность наступает очень рано; однако у всех зародышей рано или поздно клетки становятся детерминированными. Так, у зародыша морского ежа (вторичноротое) на стадии четырех бластомеров все клетки равноценны по своим потенциям к развитию. Однако у 8-клеточного и уж тем более у 16-клеточного зародыша судьба бластомеров явно предопределена.
Следует отметить, что даже у Spiralia развитие яиц нельзя считать полностью мозаичным. Некоторые детерминирующие события наступают на очень ранних стадиях, но по мере дальнейшего развития все возрастающую роль в нем начинают играть различные индукционные взаимодействия. Одна из интересных особенностей мозаичного развития состоит в том, что при этом возможно быстрое образование специализированных личинок из ограниченного числа зародышевых клеток. Такой механизм особенно выгоден для организмов, развивающиеся яйца которых взвешены в морской воде, составляя часть планктона. Индукционные взаимодействия, наблюдаемые в развитии даже типичных мозаичных зародышей, позволяют считать, что в процессе эволюции той или иной линии соотношение вкладов самодифференцировки и индукции может изменяться, особенно в тех случаях, когда у данной линии наблюдается тенденция к утрате специализированных личинок. Заключение о том, что такой процесс действительно имел место, можно сделать, рассматривая развитие асцидий, амфибий и млекопитающих - членов обширного филогенетического ряда хордовых.
Развитие асцидий носит в значительной мере мозаичный характер, как это было показано в экспериментах двух типов : 1) при сращивании двух зародышей на ранних стадиях дробления; 2) при изоляции пары бластомеров из зародыша и выращивании их в культуре. Эксперименты первого типа провел Убиш (Ubisch) в 1938 г. Сращивая зародышей попарно на стадии двух бластомеров, он обнаружил, что результаты такой операции зависят от ориентации зародышей друг относительно друга. Вообще, из сращенных зародышей развиваются двойные уроды с добавочными внутренними органами, например зародыш с одним хвостом, содержащим две хорды, каждая со своей нервной трубкой. Такой результат согласуется со способностью бластомеров дифференцироваться независимо друг от друга. Эксперименты с удалением некоторых бластомеров, которые впервые провел в широких масштабах Конклин (Е. Conklin) в 1905 г. на асцидиях, дали более определенную информацию относительно потенций отдельных бластомеров и их судьбы. На рис. 4-16 представлена карта презумптивных зачатков для 8-клеточного зародыша асцидий. Два передних бластомера анимальной половины дают эпидермис головы, присоски и головной мозг; два задних анимальных бластомера дают только эпидермис; передние бластомеры вегетативной половины дают спинной мозг, хорду и часть кишечника ; два задних вегетативных бластомера - кишечник, мезенхиму и мышцы.
Конклин хотел также выяснить, насколько строго предопределена судьба бластомеров. Выпуская 2- или 4-клеточных зародышей из пипетки, Конклин убивал один или несколько бластомеров, получая живые половинки или четвертушки зародышей; эти зародыши нормально дробились, оставаясь связанными с неделящимися убитыми бластомерами. У таких частичных зародышей развивались только те ткани, которые образовались бы из имеющихся у них живых бластомеров, если бы они находились в нормальном зародыше. Можно, конечно, возразить, что нормальному развитию частичных зародышей мешало присутствие мертвых бластомеров, но эксперименты, проведенные Ревербери (Reverberi) с изолированными бластомерами, привели к таким же результатам.
Ревербери и Минганти (Reverberi, Minganti) выращивали в культуре пары бластомеров, изолированные из 8-клеточных зародышей асцидий. Оказалось, что все эти пары обладают ограниченной способностью к дифференцировке, соответствующей их положению на составленной Конклином карте зачатков. Неожиданный результат этих экспериментов состоял в том, что, хотя кишечник, хорда, мышцы и мезенхима развивались путем самодифференцировки бластомеров, нервная ткань развиваться таким образом не могла. Изолированные передние бластомеры анимальной половины, которые должны были образовать нервную ткань, давали только эпидермис. Если же 8-клеточных зародышей расчленяли так, что передние бластомеры анимальной половины оставались в контакте с передними бластомерами вегетативной половины, предназначенными для образования хорды, то из передних анимальных бластомеров развивалась нервная ткань. Следовательно, для дифференцировки нервной ткани необходимо индукционное воздействие со стороны хорды и энтодермальных клеток. Это как бы предвосхищает гораздо более важную роль индукции, наблюдаемую у позвоночных, однако Ревербери указывает на одно существенное различие. Если во фракционированном зародыше сохраняется контакт между задними бластомерами анимальной половины, дающими эпидермис, и передними бластомерами вегетативной половины, дающими хорду, то нервная ткань не развивается. Эктодерма, образующаяся из задних анимальных бластомеров, не реагирует на нейральное индукционное влияние со стороны хорды. У амфибий, у которых индукционные взаимодействия изучены довольно подробно, наблюдается совершенно иная картина. Яйца амфибий легко получить, они крупные, хорошо выдерживают радикальные экспериментальные процедуры и поэтому широко используются в экспериментах по пересадкам ядер и целых участков зародышей. В 1925 г. Фогт (Vogt) разработал метод для определения дальнейшей судьбы разных участков зародыша у амфибий. Он установил, что, помещая на поверхность зародыша маленькие кусочки агара, пропитанные каким-нибудь прижизненным красителем, можно стабильно окрашивать небольшие группы клеток, не повреждая их при этом. Это позволяет проследить за дальнейшей судьбой окрашенных клеток и их местоположением в гаструле. На рис. 4-16 изображена карта презумптивных зачатков амфибий, на которой показаны участки ранней гаструлы, дающие затем начало нервной ткани, хорде, мезодерме и энтодерме. Относительное расположение этих участков такое же, как у зародышей асцидий, с той разницей что клетки, дающие начало мезодерме и хорде, находятся не на поверхности, как у оболочников, а лежат под слоем презумптивных энтодермальных клеток. Шпеман (Spemann) произвел реципрокные пересадки, при которых кусочек презумптивной ткани головного мозга, взятой из ранней гаструлы-донора, пересаживали в раннюю гаструлу-реципиент, в участок, дающий начало эпидермису; и наоборот, кусочек презумптивного эпидермиса пересаживали в участок зародыша-реципиента, дающий начало ткани головного мозга. Трансплантаты быстро приживлялись, и за их судьбой было нетрудно проследить, потому что донор и реципиент принадлежали к двум близкородственным видам, клетки которых четко различались по пигментации. Клетки дифференцировались в соответствии с тем участком реципиента, в который они были пересажены, т. е. их судьба не была предетерминирована.

Рис. 4-16. Карта зачатков у зародышей оболочника и лягушки. Распределение зачатков у зародыша амфибий сходно с их распределением у зародыша оболочников с той разницей, что у амфибий они располагаются в два слоя: эпидермис, энтодерма и нервная пластинка в верхнем слое, а хорда, сомиты и мезодерма под этим поверхностным слоем клеток (Ortolani, 1954 и Keller, 1975, 1976; с изменениями).
Позднее Шпеман и Мангольд (Mangold) обнаружили, что у зародыша амфибий есть один участок, способный к самодифференцировке, - спинная губа бластопора ранней гаструлы. На рис. 4-16 спинная губа изображена в виде выемки. Этот участок имеет важное значение, потому что именно здесь начинается инвагинация клеток во время гаструляции и определяется расположение дорсовентральной оси зародыша. Местоположение самой губы бластопора устанавливается вскоре после оплодотворения и определяется по появлению серого серпа. Серый серп обычно располагается против места проникновения в яйцо сперматозоида и появляется в результате акта цитоплазматической локализации (ооплазматической сегрегации), вызываемого оплодотворением, от которого зависит также и его пространственное положение. Значительная часть хорды образуется из области спинной губы. Пересаживая часть этого материала от одного зародыша другому, Шпеман и Мангольд вызывали у реципиента образование добавочного зародыша с хордой и нервной трубкой. Хорда состояла из клеток, происходивших от трансплантата, сомиты - из клеток трансплантата и реципиента, а нервная трубка - почти целиком из клеток реципиента. Таким образом основная масса клеток добавочного зародыша была образована за счет хозяина (реципиента), однако дифференцировку этих клеток в разнообразные структуры добавочного зародыша индуцировала пересаженная спинная губа.
В результате аналогичного эксперимента Мангольд и Зейдель (Mangold, Seidel) установили, что сращивание двух зародышей амфибий на двуклеточной стадии обычно приводит к развитию двойных зародышей. Серый серп, определяющий местоположение первичного организатора, на этой стадии уже существует, а поэтому очевидно, что сращенные зародыши будут содержать два независимых организационных центра, и, следовательно, у них сформируются две системы осевых органов. Очевидно, что бластомеры амфибий на ранних стадиях развития лишены той крайне сильно выраженной способности к самодифференцировке, которой обладают асцидий; однако у амфибий сохраняется мозаичная природа первичного организатора. Индукционная функция организатора имеет решающее значение для последующей дифференцировки других участков, которая зависит от цепи индукционных воздействий, инициируемых осевыми структурами.
Плацентарные млекопитающие сильно отличаются по стратегии развития как от морских беспозвоночных с их мелкими пелагическими личинками, так и от тех позвоночных, которые продуцируют крупные яйца, богатые желтком. Большинство морских беспозвоночных образуют большое число яиц, содержащих количество желтка, достаточное для обеспечения быстрого развития до стадии, на которой организм становится способным к самостоятельному питанию. В то же время у таких позвоночных, как амфибии, яиц меньше, но они очень богаты желтком, что необходимо для более длительного развития организма, способного к самостоятельному питанию. Все эти яйца содержат также запасы рибосом и мРНК, обеспечивающие быстрое развитие в начальный период, до того как в результате дробления у зародыша образуется достаточное число ядер для поддержания высокого уровня синтеза белка при участии новообразованных мРНК.
В отличие от этого у плацентарных млекопитающих яйца мелкие и содержат мало желтка или других веществ, необходимых для длительного самостоятельного синтеза белка, потому что их зародыши развиваются как бы в контейнере с питательной средой, помещенном в тело матери. Яйца млекопитающих вначале развиваются очень медленно. У зародыша мыши первые 4-5 делений дробления занимают трое суток. На четвертые сутки образуется бластоциста, состоящая примерно из 100 клеток. Имплантация происходит спустя 4,5 сут. Поскольку наличные запасы мРНК невелики, ее транскрипция в ядрах зародыша начинается у мышей очень рано - еще до первого деления дробления и имеет жизненно важное значение для осуществления ранних стадий развития.
Развитие зародышей млекопитающих, предшествующее их имплантации, приводит к образованию бластоцисты (рис. 4-17) - полой структуры, напоминающей бластулу и состоящей из клеток двух типов: клеток трофобласта, покрывающих зародыш снаружи, и внутренней клеточной массы, располагающейся в полости, ограниченной трофобластом. Из трофобласта развивается плацента, а из внутренней клеточной массы - внезародышевые оболочки и сам зародыш. Развитие зародыша до стадии бластоцисты не зависит от морфогенетической информации, получаемой в результате взаимодействия с тканями материнского организма, потому что этой стадии достигают зародыши, выращиваемые на простой питательной среде, содержащей только пируват и соли. Для раннего периода постимплантационного развития необходимы другие более сложные среды, однако результаты соответствующих экспериментов позволяют сделать вывод, что раннее постимплантационное развитие регулируется изнутри, а матка обеспечивает питание и опору; хороший обзор на эту тему составил Грэхэм (Graham, 1973).
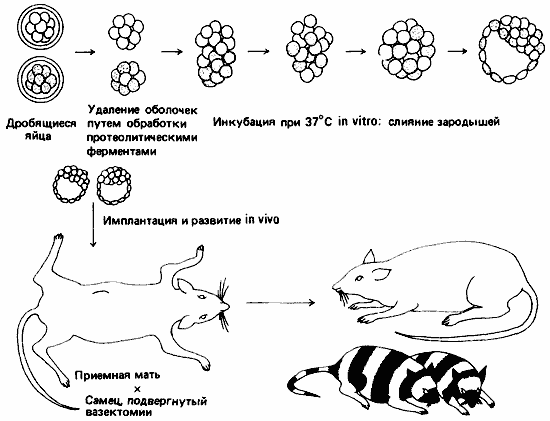
Рис. 4-17. Химерные мыши, полученные в результате сращивания зародышей двух разных генотипов на стадии дробления (Mintz, 1967).
Для того чтобы выяснить, до какой степени клетки млекопитающих способны к мозаичному развитию, были проведены эксперименты с удалением клеток и со сращиванием зародышей. Мур (N. Moore) и его сотрудники разрушали у 2-, 4- и 8-клеточных зародышей кролика все бластомеры, за исключением одного, и переносили этот последний в матку приемной матери. Из 30% таких бластомеров, взятых от 2-клеточных зародышей, 19%-от 4-клеточных, 11%-от 8-клеточных были получены нормальные крольчата. Тарковски и Вроблевска (Tarkowski, Wroblewska) разделяли бластомеры 4- и 8-клеточных зародышей мышей и выращивали их в культуре. Им удалось проследить за судьбой каждого бластомера лишь для небольшого числа диссоциированных зародышей, но некоторые полученные при этом данные оказались очень интересными. Из бластомеров одного 4-клеточного зародыша были получены три бластоцисты и один трофобластический пузырек (бластоциста, не содержащая внутренней клеточной массы). Из 8-клеточного зародыша, разделенного на пары бластомеров, были получены три бластоцисты и одна морула. Ни в одном случае не наблюдалось мозаичной дифференцировки, характерной для асцидий. Кроме того, эти результаты резко отличались от тех, которые получил Руд (Ruud) в аналогичных экспериментах с зародышами амфибий. Руд разделял бластомеры 4-клеточного зародыша и выращивал их в культуре по отдельности. Из двух бластомеров, содержавших кусочки зоны серого серпа, формировались маленькие, но полные зародыши, а два других бластомера делились, но дифференцировки не происходило.
Тарковски и Вроблевска высказали предположение, что в зародышах млекопитающих предетерминированные локализованные участки цитоплазмы не играют никакой роли. Направление дифференцировки бластомера определяется только его местоположением в ранней бластоцисте. Так, клетка, оказавшаяся снаружи, становится частью трофобласта, а клетка, попавшая внутрь, развивается во внутреннюю клеточную массу. Хильмен (Hillman) и ее сотрудники проверили это предположение, перенося меченые бластомеры во внутренние или наружные участки немеченых зародышей. Как и предсказывали Тарковски и Вроблевска, бластомеры дифференцировались в трофобласт или внутреннюю клеточную массу в соответствии со своим положением.
Зависимость судьбы клетки от ее положения и отсутствие организатора были продемонстрированы также и другим способом. Тарковски и Минц (Mintz) диссоциировали мышиные зародыши на стадии морулы и объединяли клетки двух зародышей, различающихся по генам окраски шерсти. Образовавшиеся в результате гибридные бластоцисты были имплантированы в приемную мать. Из них развились нормальные живые мышата; это были химеры, в окраске которых проявилось действие обоих генов. Схема такого эксперимента представлена на рис. 4-17.
Степень зависимости судьбы клеток млекопитающих от их местоположения в зародыше и взаимодействия с другими клетками особенно ярко продемонстрировали Минц и Илмензе (Ilmense). Эти авторы экспериментально вызывали образование тератокарцином у мышей, имплантируя в полость тела (не в матку) нормальный ранний зародыш. Развитие такого зародыша протекало беспорядочно, и он превращался в солидную опухоль, содержащую популяцию быстро делящихся стволовых клеток (эм6риокарциномных клеток), способных дифференцироваться с образованием самых разнообразных тканей. Эти солидные опухоли часто удается диссоциировать и выращивать в перитонеальной полости, получая асцитные опухоли. Асцитные опухоли состоят из эмбриональных телец, в центре которых находятся эмбриокарциномные клетки, окруженные слоем недифференцированных энтодермальных клеток. Минц и Илмензе вводили эмбриокарциномные клетки из линии асцитных опухолевых клеток, сохранявших эуплоидный набор хромосом, в бластоцисты генетически помеченной линии мышей и получали здоровых потомков, представлявших собой генетические химеры, которые были построены из нормальных тканей. происходящих как из клеток реципиента, так и из введенных ему эмбриокарциномных клеток. По-видимому, превращение тканей зародыша в тератокарциному связано с нарушением характера экспрессии генов, а не с мутационным процессом, потому что эмбриокарциномные клетки, помещенные в специализированную среду внутри бластоциста, могут дать начало нормальным тканям.
В эволюционной последовательности форм, наблюдаемой у хордовых, сохраняется один и тот же основной план строения тела, однако с течением времени роль мозаичных элементов в процессе развития постепенно становится все менее важной, пока у млекопитающих эти элементы не исчезают окончательно. И наоборот, значение индукционных взаимодействий между отдельными участками зародыша возрастает. У асцидий главное индукционное событие - образование нервной ткани под влиянием хорды. Эта фундаментальная зависимость сохраняется у эволюционно более продвинувшихся хордовых, у которых характерная для асцидий строгая самодифференцировка других частей организма сменяется системой актов детерминации, обусловленных индукционными взаимодействиями. Представляется вероятным, что в тех случаях, когда в результате возникают сходные ткани или структуры, это связано со сходными наборами экспрессирующихся генов, хотя вполне возможно, что переход от самодифференцировки к индукционным взаимодействиям сопровождается сменой триггеров, вызывающих действие генов.
В развитии хордовых помимо смягчения строго мозаичного типа развития произошло еще одно столь же важное изменение. Беррил (N. Berrill) в своей работе «Происхождение позвоночных» указал на значение изменений в соотношении между числом делений дробления, которые прошел зародыш, и началом дефинитивной дифференцировки клеток. У асцидий и оболочников вообще гаструляция начинается, как правило, между 64- и 128-клеточными стадиями. Согласно Конклину, на стадии 64 клеток у зародыша имеется 26 клеток презумптивной покровной эктодермы, 10 клеток презумптивной нервной пластинки, 4 клетки презумптивной хорды и 10 клеток мезенхимы, 4 мышечные клетки хвоста и 10 клеток презумптивной энтодермы. Некоторые из этих клеток претерпевают далее ограниченное и дискретное число клеточных делений, прежде чем приступить к окончательной дифференцировке. Так, у головастикоподобных личинок асцидий имеется 36 мышечных клеток хвоста и 40 клеток хорды. Соответственно сама эта личинка невелика.
Беррил высказал мнение, что хордовые произошли от оболочников, сохранив план строения тела их личинок в результате неотенического развития. Строгое ограничение числа клеток и общих размеров у личинок оболочников жестко ограничивало эволюционные возможности любых неотенических Prochordata. Ввиду того что размеры отдельных клеток практически ограничены, любое существенное увеличение общих размеров организма может достигаться только за счет увеличения числа клеток каждого типа. Можно соглашаться или не соглашаться с гипотезой Беррила (о неотеническом происхождении позвоночных от оболочников), поскольку палеонтологическая летопись хранит по этому поводу молчание и поскольку с равной вероятностью можно считать, что взрослые формы оболочников представляют собой специализированное терминальное добавление к жизненному циклу животных, которые первоначально во взрослом состоянии были подобны хордовым. Однако все же из табл. 4-2 видно, что среди классов хордовых имел место определенный сдвиг соотношений между числом циклов делений и сроками дифференцировки. Oikopleura - маленький неотенический оболочник, ведущий пелагический образ жизни и сохраняющий хвост во взрослом состоянии. Гаструляция у Oikopleura наступает на один цикл дробления раньше, чем у типичных оболочников-асцидий, например у Styela. Судьба клеток у обоих организмов одинакова, однако число клеток хорды и мышечных клеток хвоста показывает, что у Oikopleura детерминация происходит раньше, чем у Styela. У ланцетника Amphioxis - самого примитивного из всех настоящих хордовых - яйцо имеет такие же размеры, как и у Styela. Поскольку и гаструляция, и дифференцировка отстают у него на три цикла дробления, хорда и хвостовая мышца у личинки Amphioxis до начала питания и роста содержат в 8 раз больше клеток, чем у личинки Styela. У позвоночных Petromyzon (минога) и Triturus (тритон) эта тенденция к образованию крупных личинок зашла еще дальше. О том, что такое изменение в соотношении между числом циклов дробления и дифференцировкой имеет генетическую основу, свидетельствует существование у дрозофилы мутантного гена giant, который в гомозиготном состоянии обусловливает увеличение размеров особей (в остальном нормальных) вдвое. Такой эффект возникает в результате дополнительного цикла клеточных делений на поздних стадиях личиночной жизни.
Таблица 4-2. Зависимость между сроками клеточной детерминации у оболочников и хордовых и конечным числом клеток у их личинок (Berrill, 1955; с изменениями)
Животное Гаструляция Хорда Мышцы хвоста Диаметр яйца число дроблений примерное число клеток примерное число клеток примерное число клеток мм Oikopleura 5-6 38 20 20 0,09 Styela 6-7 76 40 36 0,13 Amphioxis 9-10 780 330 400 0,12 Petromyzon 11 2200 500 1,0 Triturus 14 16000 1200 - 2,6Если в истории развития хвоста оболочников и есть нечто поучительное, так это то, что его, вероятно, можно использовать в качестве известного обобщения, иллюстрирующего сложность эволюционных событий на ранних стадиях развития. В 1933 г. Нидхэм (J. Needham) указал, что несмотря на то что процессы развития чрезвычайно тесно и тонко интегрированы, их на самом деле можно диссоциировать - отделять друг от друга; иными словами, можно экспериментально отделить дифференцировку от роста или от клеточных делений, биохимическую дифференцировку от морфогенеза и даже отделить один от другого разные элементы морфогенеза. Значение этого высказывания для понимания эволюции огромно. Возможность такой диссоциации открывает путь к описанию последствий изменения относительных сроков различных событий, происходящих в процессе развития, для морфологической эволюции, как это сделал Гулд (Gould) в своей книге «Онтогенез и филогенез», и налагает весьма реальные ограничения на наши подходы к генетической организации процессов развития.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
6.3. Экспериментальные эволюционные машины (ЭЭМ)
6.3. Экспериментальные эволюционные машины (ЭЭМ) В предыдущем параграфе мы упоминали о том, что непрерывный рост микробных популяций в проточных системах позволяет экспериментально изучать микроэволюционные переходы, т.е. шаги эволюции.Суть непрерывного процесса
§ 25. Теории происхождения хордовых
§ 25. Теории происхождения хордовых Бытует несколько точек зрения на возникновение хордовых, которые различаются как по подходам к решению проблемы, так и по животным, избранным в качестве представителя предковых групп. Наиболее известны гипотезы возникновения хордовых
§ 26. Происхождение нервной системы хордовых
§ 26. Происхождение нервной системы хордовых Наиболее часто обсуждаемые гипотезы происхождения не могут объяснить появление одного из основных признаков хордовых — трубчатой нервной системы, которая располагается на спинной стороне тела. Мне хотелось бы использовать
4.2. Возрастные изменения морфофункциональной организации нейрона
4.2. Возрастные изменения морфофункциональной организации нейрона На ранних стадиях эмбрионального развития нервная клетка имеет большое ядро, окруженное незначительным количеством цитоплазмы. В процессе развития относительный объем ядра уменьшается. Рост аксона
3. ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА
3. ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА Строение мозга у животных разных видов разнообразно. И хотя, как следует из рис. 19.3, родственные виды, например среди ракообразных или млекопитающих, имеют общие черты в строении мозга, между собой они мало похожи. Именно с появлением
Глава 12 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЛИЧИНОЧНО-ХОРДОВЫХ КРУГЛОРОТЫХ И РЫБ
Глава 12 ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ ЛИЧИНОЧНО-ХОРДОВЫХ КРУГЛОРОТЫХ И РЫБ Высшая нервная деятельность позвоночных животных отражает одну из важных тенденций их эволюции — индивидуальное совершенствование. Эта тенденция проявляется в нарастающей продолжительности
КОГДА-ТО НАШИ ЭВОЛЮЦИОННЫЕ ПУТИ РАЗОШЛИСЬ
КОГДА-ТО НАШИ ЭВОЛЮЦИОННЫЕ ПУТИ РАЗОШЛИСЬ В области сексуальных и брачных отношений человекообразные обезьяны явно ушли от общих с человеком предков своими особыми путями. И общее у них здесь с человеком — это наличие менструальных циклов, заменивших характерную для
Эволюционные стратегии популяций
Эволюционные стратегии популяций В эволюционной экологии рассматриваются различные теоретические обобщения, показывающие межвидовые взаимоотношения. В результате взаимодействия у животных выработались собственные эволюционные стратегии. Одной из важнейших
Эволюционные изменения в организации яиц со спиральным дроблением
Эволюционные изменения в организации яиц со спиральным дроблением Для многих зародышей со спиральным дроблением, в особенности для зародышей моллюсков и кольчецов, составлены очень точные карты, на которых указана дальнейшая судьба различных клеток. Разные типы
Изменения в организации яйца при возникновении эволюционно продвинутых групп первичноротых
Изменения в организации яйца при возникновении эволюционно продвинутых групп первичноротых Среди Spiralia наблюдается несколько довольно любопытных крупных эволюционных направлений, затрагивающих ранние стадии развития. Одно из них - это сочетание невероятно
1.3. Существование самоотверженности и ее эволюционные преимущества
1.3. Существование самоотверженности и ее эволюционные преимущества Логике примитивного социал-дарвинизма противоречат факты массового героизма и самоотвержения, с существованием героической верности долгу, с самоуничтожением ради выполнения долга, с существованием
Эволюционные предпосылки гомосексуальности
Эволюционные предпосылки гомосексуальности Гомосексуальное поведение — явление отнюдь не сугубо человеческое. Оно широко распространено у животных в естественной среде обитания. Гомосексуальные отношения можно часто наблюдать в группах холостяков у павианов
Эволюционные корни ритуала
Эволюционные корни ритуала Антропологи уже давно пришли к выводу, что ритуалы в древнейших человеческих сообществах способствовали выживанию, поддерживая в данном клане или племени чувство особой идентичности и общей судьбы[91]. Ритуал постоянно напоминал членам