Эволюционные изменения в организации яиц со спиральным дроблением
Эволюционные изменения в организации яиц со спиральным дроблением
Для многих зародышей со спиральным дроблением, в особенности для зародышей моллюсков и кольчецов, составлены очень точные карты, на которых указана дальнейшая судьба различных клеток. Разные типы дробления и различия в размерах между бластомерами дают возможность проследить за судьбой отдельных бластомеров зародышей со спиральным дроблением в процессе развития. Эта их особенность в сочетании с высокомозаичным характером их развития сделала таких зародышей излюбленным объектом изучения эмбриологов.
Схема спирального дробления представлена на рис. 4-2, где изображены первые несколько делений. В результате второго деления образуются четыре бластомера - А, В, С и D. При третьем неравномерном делении образуется первый квартет микромеров. Им даны обозначения 1a, 1b, 1с и 1d, а соответствующим макромерам - 1А, 1В, 1C и 1D. Митотические веретена ориентированы таким образом, что макромеры и микромеры, возникшие в результате этого и последующих делений, расположены друг относительно друга по спирали.
При следующем делении, приводящем к 16 клеткам, микромеры первого квартета делятся равномерно, образуя 8 микромеров первого ряда. Макромеры делятся неравномерно, образуя второй квартет микромеров, обозначаемых 2а, 2b, 2с и 2d, и соответствующие макромеры. По мере дальнейшего развития как микромеры, так и макромеры продолжают делиться. Разработана специальная номенклатура для обозначения сложной системы образующихся при этом клеток, но для наших целей достаточно рассмотреть лишь общее расположение рядов микромеров при спиральном дроблении, изображенное на рис. 4-11, I. На этой упрощенной схеме показано только по четыре клетки каждого ряда. У настоящего зародыша в некоторых рядах будет, конечно, больше чем по четыре клетки вследствие продолжающегося дробления микромеров. Как на этой, так и на других сходных схемах, приведенных в данном разделе, многие детали опущены и рассматриваемые зародыши изображены лишь в общих чертах.
На рис. 4-11 показана типичная судьба различных бластомеров у зародышей Spiralia. Первые три ряда микромеров (не заштрихованы) образуют эктодерму зародыша и некоторые эктодермальные структуры; клетка, покрытая точками (4d), дает начало мезодерме; из макромеров (заштрихованы) развивается энтодерма - презумптивная средняя кишка. Подобный набор клеточных линий удивительно постоянен для всех зародышей Spiralia: у многоветвистых плоских червей, у кольчецов, а также у брюхоногих и двустворчатых моллюсков спиральное дробление протекает в основном одинаково, и судьбу клеток в этих группах можно непосредственно сопоставлять.
Несмотря на такое постоянство характера дробления у зародышей Spiralia, между ними имеются некоторые существенные различия, вскрывающие эволюционные модификации в способности к самодифференцировке бластомеров у зародышей с мозаичным дроблением, принадлежащих к разным группам. Это: 1) изменения характера локализации, в результате которых судьба определенного участка или бластомера изменяется по сравнению с их судьбой у предковых форм; 2) изменения относительных скоростей клеточного деления, приводящие к модификациям относительных размеров клеток и их числа; 3) изменения цитоскелетного матрикса, приводящие к изменению местоположения митотического аппарата во время дробления, что в свою очередь приводит к сдвигу пропорций или распределения бластомеров у данного зародыша.
Можно показать, что в рамках основной модели развития, характерной для Spiralia, встречаются модификации всех этих трех типов. Изменения в судьбе клеток могут быть очень незначительными, как в случае возникновения эктомезодермы, из которой развиваются такие мезодермальные структуры, как личиночная мышца. Например, на рис. 4-11, II и III показано происхождение эктомезодермы у двух моллюсков - брюхоногого Crepidula и двустворчатого Unio. У Crepidula эктомезодерма образуется из трех микромеров второго квартета, а у Unio - только из одного микромера.
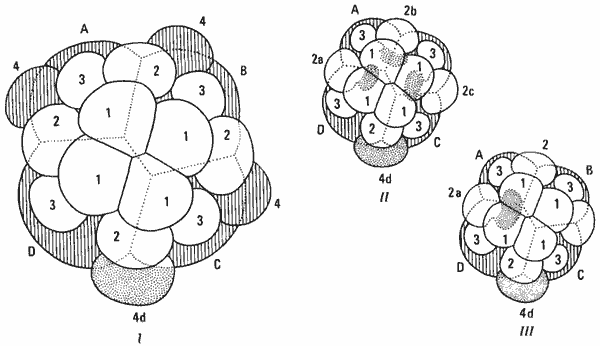
Рис. 4-11. Схематическое изображение характера дробления и судьбы различных клеток у зародышей Spiralia. I. Обобщенная схема дробления зародышей Spiralia; показаны первые три квартета микромеров (не заштрихованы), дающие начало зародышевой эктодерме, клетки 4d (пунктир), дающие начало мезодерме, и макромеры (заштрихованы), дающие начало энтодерме. II и III. Схемы спирального дробления у зародышей Crepidula (II) и Unio (III). У этих зародышей дробление протекает сходным образом, но различается по месту возникновения эмбриональной эктомезодермы из второго квартета микромеров (Wilson, 1898).
У кольчецов эволюция олигохет из полихет сопровождалась рядом резко выраженных изменений в судьбе различных клеток. В развитии полихет имеется высокодифференцированная личиночная стадия - трохофора (рис. 4-12, III). Для того чтобы могли образоваться весьма сложные личиночные органы, многие бластомеры зародыша должны дать начало временным органам личинок, тогда как остальные бластомеры образуют недифференцированные зачатки дефинитивных органов. На рис. 4-12, I изображен 40-клеточный зародыш многощетинкового червя Podarke. Заштрихованные участки зародыша соответствуют заштрихованным участкам на карте презумптивных зачатков для бластулы Podarke (рис. 4-12, II). Большая часть показанных на схемах участков соответствует определенным структурам, специфичным для трохофоры, таким как теменной султан и прототрох. При метаморфозе некоторые личиночные ткани, например личиночная мышца и прототрох, подвергаются гистолизу и исчезают, тогда как другие личиночные ткани, такие как первичный рот и средняя кишка, превращаются в эквивалентные структуры взрослых особей. Зачатки этих структур начинают дифференцироваться. Так мезодермальные тяжи дают начало туловищным сомитам.
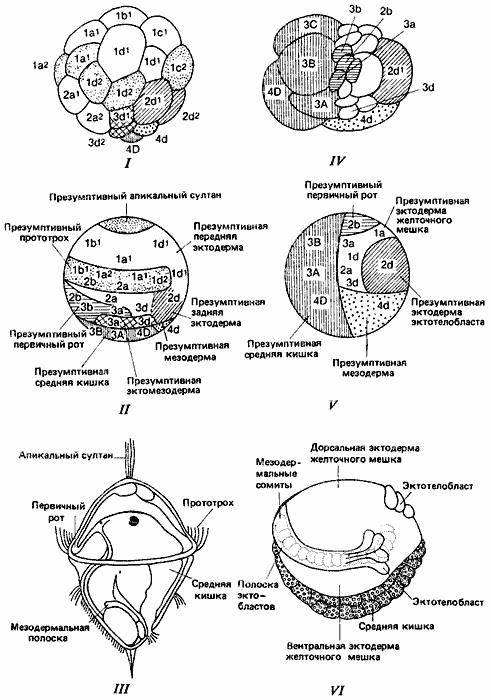
Рис. 4-12. Сравнение развития у двух кольчатых червей - полихеты Podarke и олигохеты Tiibifex. I. Зародыш Podarke на стадии 40 клеток. II. Карта зачатков Podarke на стадии бластулы. III. Трохофора Podarke. IV. Дробящийся зародыш Tubifex. V. Карта зачатков Tubifex. VI. Гаструляция у Tubifex, непосредственно ведущая к развитию сегментированной взрослой особи. (Рис. I - Treadwell, 1901; рис. II-VI - Anderson, 1973.)
В отличие от этого в тех случаях, когда образование личиночных структур в процессе эволюции подавляется, большая часть бластомеров зародыша непосредственно развивается в структуры взрослого организма. Так обстояло дело в процессе эволюции олигохет, в развитии которых нет высокодифференцированной личиночной стадии и у которых развитие сегментированного тела взрослой особи начинается во время гаструляции. На рис. 4-12, VI показана гаструляция у олигохеты - трубочника Tubifex. У ее зародыша не происходит образования обширного участка эктодермы на переднем конце тела (теменной пластинки) с отходящим от него прототрохом. Когда средняя кишка инвагинирует, у зародыша начинают формироваться сомиты, превращающиеся в дальнейшем в сегменты тела взрослого червя. Это резко отличается от процесса развития у полихет, у личинок которых при вполне сформированных первичном рте и средней кишке презумптивные сомиты представлены в виде недифференцированных зачатков. Возникающие в результате этого изменения дальнейшей судьбы клеток зародыша показаны на рис. 4-12, IV-VI, на которых стадии развития Tubifex сопоставлены с соответствующими стадиями развития полихет. Как можно видеть на рис. 4-12, IV, хотя у Tubifex дробление все еще происходит по спиральному типу, оно, однако, значительно отличается от дробления у полихет. Микромеры, соответствующие тем, которые у полихет дают начало передней эктодерме (теменной пластинке), составляют относительно гораздо меньшую часть общей массы зародыша, тогда как относительные размеры клеток, дающих презумптивные среднюю кишку и мезодерму, значительно возросли.
Судьба клеток у зародыша Tubifex также изменилась. Как можно видеть, сравнивая карту презумптивных зачатков, изображенную на рис. 4-12, V с аналогичной картой для полихет (рис. 4-12, II), хотя презумптивные участки на обеих картах в общем сходны, но у зародыша Tubifex обнаруживается много специфичных изменений. Презумптивные участки эктомезодермы, личиночной эктодермы, теменного султана и прототроха утрачены. Те бластомеры, из которых у полихет развиваются эти структуры, у олигохет образуют другую эктодермальную структуру - желточный мешок. Другие участки, такие как клетка 4d, дающая в обеих группах мезодерму взрослой особи, имеют такую же конечную судьбу, но изменились пути развития, по которым эти участки к ней приходят. Однако, как показал Пеннерс (Penners), у Tubifex сохранился строго мозаичный тип развития, характерный для кольчецов. По-видимому, локализованные детерминанты изменились и вызывают у Tubifex генную экспрессию иного типа, нежели у предковых полихет. Дальнейшую модификацию можно наблюдать в развитии пиявки Erpobdella, протекающем в питательном коконе. Здесь бластомеры 1 А, 1В и 1C не делятся; их обгоняют в росте другие клетки, и они начинают функционировать как альбуминотрофные клетки, а большую часть тела взрослой особи составляют клетки, происходящие из бластомера 1D.
Эволюционные изменения в судьбе клеток, описанные здесь для кольчецов, не ограничиваются модификациями локализованных индукторов дифференцировки; они требуют также изменения относительных сроков дробления разных бластомеров. Так, макромеры, которые у полихет делятся несколько раз, чтобы дать начало микромерам, участвующим в образовании таких зародышевых структур, как прототрох или эктомезодерма, у пиявок не делятся вовсе. Конечно, изменения относительных сроков клеточных делений, имевшие эволюционное значение, происходили не только у кольчецов; они ясно видны также у моллюсков.
У пресноводных Unionidae, относящихся к двустворчатым моллюскам, в остальном довольно ординарных, личинки в процессе эволюции выработали совершенно необычный образ жизни. Unionidae живут в проточных водах; взрослые особи прикреплены к субстрату, а свободноплавающих личинок течение безжалостно сносило бы вниз. Репродуцирующиеся особи производят многочисленных зародышей, развивающихся в родительском организме до достижения личиночной стадии - так называемого глохидия. Глохидий имеет вид миниатюрного медвежьего капкана (рис. 4-13, I). Для завершения развития эта личинка должна прикрепиться к жабрам или плавникам какой-нибудь рыбы. Здесь она в течение нескольких недель ведет паразитический образ жизни, после чего отпадает и продолжает свое существование на дне как обычный двустворчатый моллюск.
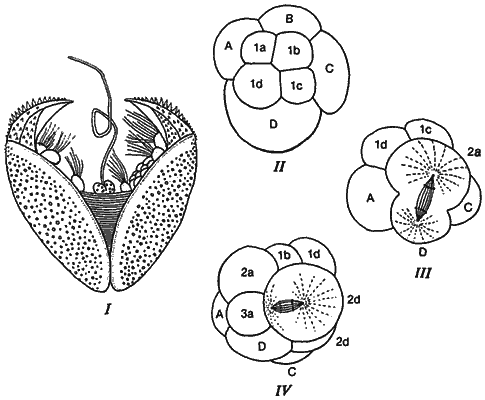
Рис. 4-13. Зародыши и личинки двустворчатого моллюска Unio. I. Глохидий с его сенсорными волосками и со створками, напоминающими медвежий капкан в миниатюре. II. Зародыш на стадии 8 бластомеров с типичным спиральным дроблением. III. Образование относительно крупного микромера 2d, дающего раковинную железу личинки. IV. Дальнейшее дробление с образованием крупного микромера 2а, дающего замыкательную мышцу личинки (Lillie, 1895).
У глохидия имеются чувствительные сенсорные волоски, и при малейшей опасности створки его раковинки, снабженные мощными крючками, захлопываются. У некоторых видов глохидии просто лежат на дне в надежде на то, что какая-нибудь рыба случайно проплывет мимо. У других видов у самки имеется мантия, край которой модифицирован, напоминая глаза и тело гольяна. Когда Unio готова выпустить наружу личинок, край мантии начинает волнообразно изгибаться, предположительно для того, чтобы привлечь внимание проплывающих мимо рыб к «гольяну» и заставить подходящего хозяина подойти достаточно близко к только что выпущенным в воду глохидиям.
В своей работе по ранним стадиям развития пресноводной Unio, опубликованной в 1898 г., Лилли (Lillie) убедительно показал, что дробление у этого моллюска, сохраняя основные черты, типичные для Spiralia, претерпело в отношении скорости и общего характера изменения, связанные с необходимостью формирования специализированных личиночных структур. У большинства моллюсков и кольчецов микромеры первого ряда образуют теменную пластинку и прототрох. У личинок Unio, у которых дробление первого ряда микромеров происходит медленнее, чем дробление второго их ряда, эти структуры отсутствуют. Такое замедление выявляется при сравнении распределения клеток, имеющихся на стадии 32 бластомеров у Unio и у «идеализированного» зародыша со спиральным дроблением (табл. 4-1). Второй ряд микромеров дает начало большей части личиночных структур (и массы) личинок Unio. Кроме того, некоторые зародышевые структуры у глохидия очень велики по сравнению с другими его частями. Одна из таких структур - раковинная железа, которая вырабатывает относительно массивную раковину зародыша. Эта железа образуется всего из одного микромера второго ряда - микромера 2d. Дробление несколько модифицируется, так что микромер 2d оказывается крупнее своего сестринского 2D-макромера и фактически является самой крупной клеткой зародыша. На рис. 4-13, II-IV изображены стадии дробления зародыша Unio. У 8-клеточного зародыша (рис. 4-13, II) макромеры и микромеры распределены типичным для Spiralia образом. По мере дальнейшего дробления зародыша, как это показано на рис. 4-13, III (вид сбоку), макромер D делится, образуя крупную клетку 2d. Эта клетка продолжает делиться, образуя ряд мелких клеток (рис. 4-13, IV). Огромные относительные размеры бластомера 2d у Unio можно оценить, сравнивая рис. 4-13, IV и 4-12, I. Вторая крупная клетка второго квартета микромеров - это бластомер 2а (также показанный на рис. 4-13, IV), дающий начало личиночной эктомезодерме, из которой развивается крупная замыкающая мышца, обеспечивающая захлопывание раковины глохидия. Клетки, получающиеся в результате деления клеток 2а и 2d, также делятся быстрее, чем другие микромеры второго квартета.
Таблица 4-1. Относительные скорости дробления у Unio по сравнению с «идеализированным» зародышем со спиральным развитием на стадии 32 бластомеров (Lillie, 1898; с изменениями)
Клетки Идеальный зародыш Зародыш Unio Первый квартет микромеров 16 10 Второй квартет микромеров 8 13 Третий квартет микромеров 4 4 Клетка 4d (презумптивная мезодерма) 1 Макромеры 4 4 Общее число бластомеров 32 32Лилли ясно представлял себе, что в этих модификациях спирального дробления у моллюсков участвует несколько факторов. Хотя судьба клеток не изменяется в общем смысле, т. е. в отношении того, из каких бластомеров образуется эктодерма, мезодерма и энтодерма, личиночная форма, как мы это видели среди кольчецов у олигохет, значительно модифицировалась по сравнению с предковой трохофорой. В число необходимых адаптации, затрагивающих дробление, входят изменения относительных скоростей клеточного деления и относительных размеров бластомеров. Последнее достигается в результате модификаций цитоскелетного матрикса, контролирующего местоположение митотических веретен.
Локализация веретена определяет не только правое и левое направление дробления, как это видно у Limnaea, но также относительные размеры дочерних клеток. Если веретено располагается в центре клетки, то борозда дробления, образующаяся в плоскости метафазной пластинки, проходит по экватору, и в результате дробления получаются две одинаковые дочерние клетки. Если же, однако, веретено находится на значительном расстоянии от центра, то борозда дробления также смещена и одна из дочерних клеток оказывается гораздо крупнее другой. Это хорошо видно на рис. 4-13, III, где в результате деления клетки D образуются две дочерние клетки, сильно различающиеся по величине.
Регуляция скорости дробления также связана с цитоплазмой зародышей. В 1904 г. Вилсон (Е. Wilson) установил, что в 16-клеточном зародыше Patella четыре клетки, составляющие первый квартет микромеров, уже детерминированы как первичные трохобласты и дифференцируются в 16 ресничных клеток прототроха. По мере продолжения дробления каждый из этих первичных трохобластов делится еще дважды, а затем за 10 ч у него вырастают реснички, расположенные поперечными рядами. Вилсон сумел изолировать отдельные первичные трохобласты из 16-клеточного зародыша. Изолированные трохобласты, как и в норме, делились еще два раза, а затем прекращали деление, и примерно на 10-м часу у них развивались реснички, расположенные обычным образом. Итак, у этих зародышей мозаичный характер развития проявился не только в дифференциальной способности изолированных трохобластов к образованию специфически расположенных ресничных клеток, но также и в регуляции скорости и числа клеточных делений.
Данные в пользу того, что такая регуляция сроков обусловлена действием генов во время оогенеза, получены при изучении гибридов от скрещиваний между видами, различающимися по скорости развития. Такие гибриды обычно развиваются со скоростью, характерной для материнского вида: отцовские же признаки проявляются у них на сравнительно поздних стадиях развития. Например, лягушки Rana pipiens и R. palustris заметно различаются по скорости дробления, но гибриды между ними развиваются и достигают стадии взрослых животных. Как показал Мур (Moore), клеточное деление при дроблении происходит у них с такой скоростью, как у материнского вида. Подобным же образом у гибридов между морскими ежами Paracentrotus lividus и Arbacia lixula, как установили Уитли и Болцер (Whiteley, Baltzer), скорость дробления соответствует таковой у материнского вида. Еще одно, особенно наглядное, доказательство регуляции сроков развития цитоплазмой яйца было получено в экспериментах Минганти (Minganti). Он проводил опыты по оплодотворению энуклеированных яиц асцидии Ascidia malaca спермой другой асцидии Phallusia mamillata. Значительная доля получавшихся при этом зародышей достигала личиночных стадий, и, хотя эти зародыши содержали только один геном из отцовского вида (Phallusia), скорость развития (которая у этих двух видов различна) соответствовала скорости развития материнского вида (Ascidia). Таким образом, факторы, регулирующие скорость дробления, могут обособляться в определенных бластомерах таким же образом, как и факторы, определяющие специфичные типы дифференцировки.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
6.3. Экспериментальные эволюционные машины (ЭЭМ)
6.3. Экспериментальные эволюционные машины (ЭЭМ) В предыдущем параграфе мы упоминали о том, что непрерывный рост микробных популяций в проточных системах позволяет экспериментально изучать микроэволюционные переходы, т.е. шаги эволюции.Суть непрерывного процесса
4.2. Возрастные изменения морфофункциональной организации нейрона
4.2. Возрастные изменения морфофункциональной организации нейрона На ранних стадиях эмбрионального развития нервная клетка имеет большое ядро, окруженное незначительным количеством цитоплазмы. В процессе развития относительный объем ядра уменьшается. Рост аксона
3. ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА
3. ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА Строение мозга у животных разных видов разнообразно. И хотя, как следует из рис. 19.3, родственные виды, например среди ракообразных или млекопитающих, имеют общие черты в строении мозга, между собой они мало похожи. Именно с появлением
КОГДА-ТО НАШИ ЭВОЛЮЦИОННЫЕ ПУТИ РАЗОШЛИСЬ
КОГДА-ТО НАШИ ЭВОЛЮЦИОННЫЕ ПУТИ РАЗОШЛИСЬ В области сексуальных и брачных отношений человекообразные обезьяны явно ушли от общих с человеком предков своими особыми путями. И общее у них здесь с человеком — это наличие менструальных циклов, заменивших характерную для
Эволюционные истоки морфологических особенностей человека
Эволюционные истоки морфологических особенностей человека Как с эволюционной точки зрения интерпретировать переход к прямохождению, исчезновение волос на теле, нетипичное для обезьян распределение жировой ткани, стремительное увеличение размеров головного мозга и
Эволюционные стратегии популяций
Эволюционные стратегии популяций В эволюционной экологии рассматриваются различные теоретические обобщения, показывающие межвидовые взаимоотношения. В результате взаимодействия у животных выработались собственные эволюционные стратегии. Одной из важнейших
Разрывы, недостающие звенья и эволюционные механизмы
Разрывы, недостающие звенья и эволюционные механизмы Отсутствие предковых форм или форм, промежуточных между отдельными ископаемыми видами, не является некой странной особенностью ранней истории Metazoa. Разрывы представляют собой общее явление и встречаются по всей
Изменения в организации яйца при возникновении эволюционно продвинутых групп первичноротых
Изменения в организации яйца при возникновении эволюционно продвинутых групп первичноротых Среди Spiralia наблюдается несколько довольно любопытных крупных эволюционных направлений, затрагивающих ранние стадии развития. Одно из них - это сочетание невероятно
Эволюционные изменения в организации яиц хордовых
Эволюционные изменения в организации яиц хордовых Для Spiralia характерно развитие, при котором отдельные бластомеры на ранних стадиях дробления уже запрограммированы к дифференцировке в определенном направлении, не изменяющемся даже в том случае, если изолировать их от
1. ЕСТЕСТВЕННАЯ АГРЕССИВНОСТЬ И ЭВОЛЮЦИОННЫЕ МЕХАНИЗМЫ, ЕЕ ОГРАНИЧИВАЮЩИЕ
1. ЕСТЕСТВЕННАЯ АГРЕССИВНОСТЬ И ЭВОЛЮЦИОННЫЕ МЕХАНИЗМЫ, ЕЕ ОГРАНИЧИВАЮЩИЕ Три закона робототехники1. Робот не может причинить вред человеку или своим бездействием допустить, чтобы человеку был причинен вред.2. Робот должен повиноваться всем приказам, которые отдает
1.3. Существование самоотверженности и ее эволюционные преимущества
1.3. Существование самоотверженности и ее эволюционные преимущества Логике примитивного социал-дарвинизма противоречат факты массового героизма и самоотвержения, с существованием героической верности долгу, с самоуничтожением ради выполнения долга, с существованием
Когда-то наши эволюционные пути разошлись
Когда-то наши эволюционные пути разошлись Человекообразные в области сексуальных и брачных отношений явно ушли от общих с человеком предков своими особыми путями. Общее с человеком у них только наличие менструальных циклов, заменивших овуляцию раз в год, характерную
1.5. Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера
1.5. Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера С деятельностью живых систем связана та часть поверхности Земли, которая объединена под названием биосферы. Биосфера, являющаяся самой крупной экосистемой, представляет собой
Эволюционные предпосылки гомосексуальности
Эволюционные предпосылки гомосексуальности Гомосексуальное поведение — явление отнюдь не сугубо человеческое. Оно широко распространено у животных в естественной среде обитания. Гомосексуальные отношения можно часто наблюдать в группах холостяков у павианов
Эволюционные корни ритуала
Эволюционные корни ритуала Антропологи уже давно пришли к выводу, что ритуалы в древнейших человеческих сообществах способствовали выживанию, поддерживая в данном клане или племени чувство особой идентичности и общей судьбы[91]. Ритуал постоянно напоминал членам