Природа локализованных информационных молекул и их действие
Природа локализованных информационных молекул и их действие
Представляется вероятным, что локализованные цитоплазматические молекулы, модифицирующие экспрессию генов на ранних стадиях развития специфичным для каждого участка образом, отличаются разнообразием. Локализованные детерминанты встречаются у животных, относящихся ко многим типам, в том числе у гребневиков, немертин, кольчецов, моллюсков, членистоногих, иглокожих, оболочников и хордовых. В некоторых случаях действие локализованных детерминантов проявляется уже при первом делении яйца, как, например, в виде крупных полярных лопастей у ряда представителей Spiralia, в том числе у ничем другим непримечательной улитки Ilyanassa.
Последовательность событий, происходящих при первом митотическом делении оплодотворенного яйца Ilyanassa, показана на рис. 4-5. Вскоре после дробления на богатом желтком вегетативном полюсе яйца (расположенном против анимального полюса, на котором появляется борозда дробления) возникает выступающее наружу вздутие цитоплазмы, называемое полярной лопастью. Эта лопасть располагается перпендикулярно оси митотического веретена. Важно указать, что лопасть не содержит ядра и состоит только из цитоплазмы. Когда борозда дробления начинает углубляться, шейка, соединяющая лопасть с зародышем, быстро сжимается, превращаясь в тонкую нить цитоплазмы. По окончании делений дробления шейка полярной лопасти быстро увеличивается в диаметре и лопасть втягивается одним из бластомеров. Клетку, в которую включается лопасть, обозначают как бластомер CD, а другую клетку - как бластомер АВ.
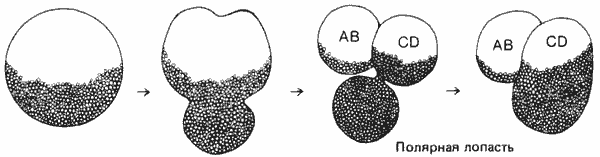
Рис. 4-5. Появление и втягивание безъядерной лопасти при первом делении дробления у зародыша брюхоногого моллюска Ilyanassa (рисунок с натуры). В бластомерах АВ и CD имеются ядра, а богатая желтком полярная лопасть лишена ядра.
Полярную лопасть легко удалить, и зародыши, которые ее лишены, продолжают развиваться с такой же скоростью, как и нормальные зародыши. Однако из нормальных зародышей развивается сложная личинка, называемая велигером (парусником), у которой имеются разнообразные структуры, тогда как зародыши, лишенные полярной лопасти, образуют всего лишь комочек клеток, покрытый ресничками (рис. 4-6). Такой эффект возникает не просто в результате удаления некоторой части массы зародыша или сокращения поступления питательных веществ. Если поставить другой эксперимент - отделить на той же двуклеточной стадии бластомер АВ от бластомера CD, то из бластомера АВ, так же как из зародыша, лишенного полярной лопасти, не развивается полностью дифференцированный велигер. Однако из бластомера CD, обладающего примерно такой же массой, как лишенный лопасти зародыш, получается нормальный, хотя и маленький, велигер.
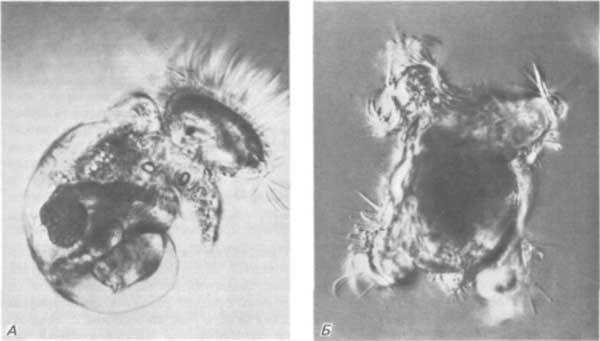
Рис. 4-6. Функция полярной лопасти в развитии. А. Нормальная личинка-велигер, имеющая глаза, ногу, раковину и внутренние органы. Б. Зародыш на той же стадии развития, у которого при первом дроблении была удалена полярная лопасть; никаких организованных структур у него нет. (Фотография любезно предоставлена Newrock.)
Возможно, что в полярной лопасти Ilyanassa заключены какие-то специфичные мРНК. Ньюрок и Рэфф (Newrock, Raff) установили, что у лишенных лопасти и у нормальных зародышей синтез белка даже на стадиях, предшествующих началу морфогенеза, протекает по-разному. Эти различия в синтезе белка наблюдались также между лишенными лопасти и нормальными зародышами при непрерывном выращивании их в присутствии таких количеств актиномицина, которые полностью подавляют синтез РНК. Эти результаты были интерпретированы как указание на то, что в полярной лопасти обособляются какие-то преформированные типы мРНК, потому что в зародышах, выращиваемых в присутствии актиномицина, происходит трансляция только тех мРНК, которые уже содержались в цитоплазме яйца ко времени формирования этой лопасти. Однако Брандхорст и Ньюрок (Brandhorst, Newrock), используя двумерный гель-электрофорез, позволяющий выявить несколько сот наиболее часто встречающихся видов белка, не обнаружили никаких качественных различий между белками нормальных зародышей и зародышей, лишенных полярной лопасти. Они, однако, обнаружили резко выраженные количественные различия, которые, возможно, и привели к результатам, полученным Ньюроком и Рэффом. Ни в той ни в другой работе не удалось выявить потенциально очень большое число редких видов мРНК, которые могли быть дифференциально обособлены в полярной лопасти.
Со сходной проблемой пришлось столкнуться при изучении зародышей морских ежей. При четвертом делении дробления у этих животных образуются клетки трех типов - мезомеры, макромеры и микромеры, значительно различающиеся по своим размерам. Клеткам этих трех типов уже в момент возникновения уготованы различные и вполне определенные судьбы. Роджерс и Гросс (Rodgers, Gross), а также Эрнст (Ernst) и ее сотрудники, используя метод гибридизации нуклеиновых кислот, обнаружили, что высокоповторяющаяся РНК распределена между этими тремя типами клеток неравномерно. Подобным же образом Мидзуно (Mizuno et al.) и Уайтли (Whiteley et al.) сообщают, что у разных бластомеров различны преобладающие транскрипты повторяющихся последовательностей ДНК. Картина осложняется наблюдением Туфаро и Брандхорста (Tufaro, Brandhorst) об отсутствии между бластомерами различий по характеру синтеза примерно 1000 видов белков, разделяемых методом двумерного гель-электрофореза. Локализованные последовательности, выявленные Роджерсом и Гроссом, а также Эрнст и ее сотрудниками, возможно, слишком редки, чтобы продуцировать достаточные количества белка, которые можно выявить методом двумерного гель-электрофореза, или это могут быть те последовательности, которые не входят в состав мРНК. Самые определенные доказательства того, что мРНК могут служить регуляторами морфогенеза, дает работа лаборатории Калтхофа (Kalthoff et al.) на яйцах двукрылого Smittia. У насекомых передний и задний концы яйца детерминируются во время оогенеза. В процессе нормального развития на переднем конце тела образуется голова и три грудных сегмента, а на заднем - ряд брюшных сегментов (и зачатковых клеток). В 1968 г. Калтхоф и Сандер сообщили, что облучение цитоплазмы на переднем конце яйца ультрафиолетом приводит к развитию зародыша, у которого вместо головы, груди и передних брюшных сегментов образуется как бы в зеркальном отображении второй задний конец тела. На рис. 4-7 изображены нормальный зародыш и урод с дуплицированным брюшком. Существуют две группы данных, указывающие на то, что развитие переднего конца тела детерминирует РНК. Одна группа данных получена в экспериментах с инактивацией РНК ультрафиолетом. В спектре действия ультрафиолета имеются пики при 265 и 285 нм, соответствующие максимальному эффекту; это позволяет считать, что ультрафиолет оказывает свое действие на комплекс нуклеиновая кислота-белок. Интересно отметить, что эффекты облучения ультрафиолетом обратимы; это достигается последующим воздействием на облученное яйцо света с длиной волны 320-480 нм. Ультрафиолет индуцирует в нуклеиновых кислотах образование пиримидиновых димеров и инактивирует эти молекулы. Обратимость этого эффекта под действием света происходит под влиянием фоточувствительного фермента, обусловливающего репарацию пиримидиновых димеров.
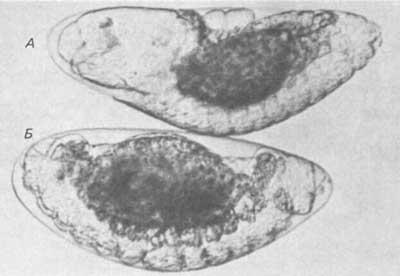
Рис. 4-7. Нормальные зародыши и зародыши с двойным брюшком двукрылого Smittia. У нормального зародыша (А) видна слева развивающаяся голова, а справа-брюшные сегменты. У зародыша Б, которого облучали ультрафиолетом, голова отсутствует и на обоих концах тела развиваются брюшные сегменты (Kalthoff, 1969).
Вторая группа данных, указывающих на детерминирующую роль РНК в развитии переднего конца тела зародыша, получена в результате непосредственного воздействия специфических ферментов на цитоплазму переднего конца зародыша. Кандлер-Зингер (Kandler-Singer) и Калтхоф погружали зародыши в среду, содержащую исследуемый фермент, и затем прокалывали их в определенных местах. Образование дуплицированного брюшка происходило лишь в тех случаях, когда в передний конец яйца проникала активная РНКаза. Если использовалась неактивная РНКаза или если она проникала не в передний конец яйца, а в другие его участки, то дуплицированное брюшко получалось лишь в очень немногих случаях.
Есть и другие более косвенные данные о том, что РНК играет роль детерминанта и у других организмов. Образование зародышевых клеток на заднем конце яиц насекомых (и многих других организмов) зависит от детерминантов, называемых полярными гранулами, которые легко наблюдать в микроскоп. В ряде интересных экспериментов Илмензе и Маховалд (Ilmensee, Mahowald) вводили цитоплазму из заднего конца яйца дрозофилы одной генетически меченной линии в передний конец яйца другой генетически меченной линии. При этом на переднем конце яйца формировались зародышевые клетки. Полярные гранулы, подобно детерминантам переднего конца тела, чувствительны к облучению ультрафиолетом и, судя по реакции на цитологические красигели, содержат большое количество РНК. Домен и Вердонк (Dohmen, Verdonk) обнаружили в полярных лопастях зародышей некоторых брюхоногих моллюсков структуры, аналогичные полярным гранулам. Эти структуры, которые, как было установлено при помощи специфичных красителей, богаты РНК, по-видимому, содержат детерминанты, специфичные для полярных лопастей. Было бы заманчиво попытаться установить связь между этими структурами и очевидным обособлением мРНК в полярных лопастях, которое наблюдали Ньюрок и Рэфф, но достаточных оснований для этого у нас нет.
Возможно, из-за того что в настоящее время все внимание молекулярной биологии сосредоточено на нуклеиновых кислотах, экспериментальным изучением белков как локализованных детерминантов ядерной активности занимаются мало. А между тем такие белковые молекулы почти несомненно существуют. Так, у аксолотля имеется мутация, при которой отсутствие одного определенного белка оказывает резко выраженное воздействие на развитие. Эта мутация, обозначаемая буквой о (oocyte deficient), приводит к тому, что яйца, отложенные самками, гомозиготными по мутантному аллелю о, прекращают дробление и гибнут примерно в то время, когда нормальные зародыши проходят гаструляцию. Мутация о - классическая мутация с материнским эффектом, т. е. развитие потомков зависит только от генотипа матери (см. гл. 7). Таким образом яйца, отложенные самкой, гомозиготной по аллелю о (о/о), не развиваются даже при оплодотворении нормальной спермой. В отличие от этого все яйца гетерозиготной самки (о/ + ), даже та их половина, которая несет аллель о, развиваются нормально. Поскольку генотип самца не играет роли, самок, имеющих генотип о/о, можно получить, оплодотворяя яйца гетерозиготных самок спермой самцов, несущих аллель о. Бриггс и Кассенс (Briggs, Cassens) обнаружили, что неспособность к развитию яиц, отложенных самками о/о, можно преодолеть, если вскоре после оплодотворения ввести в них цитоплазму нормальных яиц. Результаты, полученные Бриггсом и Джастусом (Justus), показывают, что корректирующий фактор, отсутствующий в неполноценных яйцах, представляет собой белок.
В ряде исследований, в которых в яйца вводили белки, было твердо установлено, что некоторые белки легко проникают в ядра. Такие белки могут оказывать влияние на поведение ядра, как это обнаружили Бенбау и Форд (Benbow, Ford), которым удалось индуцировать синтез ДНК в ядрах, обрабатывая изолированные ядра лягушек белком, выделенным из цитоплазмы яиц и зародышей.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Тропическая природа
Тропическая природа
ГМ растения с заданным химическим составом и структурой молекул (аминокислоты, белки, углеводы)
ГМ растения с заданным химическим составом и структурой молекул (аминокислоты, белки, углеводы) Основной закон рационального питания диктует необходимость соответствия уровней поступления и расхода энергии. Уменьшение энерготрат современного человека ведет к
ЧЕЛОВЕК И ПРИРОДА
ЧЕЛОВЕК И ПРИРОДА Выше мы говорили о больших проблемах, связанных с охраной первобытной природы в современном мире. Попытаемся теперь перейти к «синтезу» этих проблем, которые на первый взгляд мало совместимы, и попробуем развить философию сохранения природы и ее
Изучение молекул позволяет распутать эволюционный клубок
Изучение молекул позволяет распутать эволюционный клубок Метод гибридизации ДНКСтараясь установить, насколько тесно родство между отдельными видами, ученые разработали для достижения этой цели три основных метода, опирающихся на измерение степени различий в ДНК
Природа и люди
Природа и люди Тихое, мелодичное жужжание все лето не умолкает в углу лаборатории. Здесь, торцом приставленный к подоконнику, стоит плоский стеклянный улей, в котором живет небольшая семья пчел. Ее поселяют сюда весной, перед тем как зацветают сады, и пчелы скоро
Концы молекул ДНК — объект повышенного внимания
Концы молекул ДНК — объект повышенного внимания Устройство концов хромосом давно интересовало исследователей. Теперь стало ясно, что они устроены по особому. И эту некодирующую белки ДНК никак нельзя назвать «эгоистичной». Концевой участок ДНК, называемый теломерным,
Концы молекул ДНК — объект повышенного внимания
Концы молекул ДНК — объект повышенного внимания Устройство концов хромосом давно интересовало исследователей. Теперь стало ясно, что они устроены по особому. И эту некодирующую белки ДНК никак нельзя назвать «эгоистичной». Концевой участок ДНК, называемый теломерным,
1. Человеческая природа
1. Человеческая природа «Откуда мы пришли? Кто мы? Куда мы идем?» — эти предельно простые вопросы, написанные Полем Гогеном в углу его гениального таитянского холста, на самом деле являются центральными вопросами религии и философии. Сможем ли мы ответить на них? Иногда
Наследственность, или «природа»
Наследственность, или «природа» Если у высокого отца рослый сын, принято говорить, что сын унаследовал рост отца. Как обстоит дело в действительности, мы увидим ниже. Что же касается наследственного кретинизма, то здесь мы сталкиваемся с особым случаем: родители карликов
ТАЙНЫ МОЛЕКУЛ
ТАЙНЫ МОЛЕКУЛ
Природа и люди
Природа и люди Тихое, мелодичное жужжание все лето не умолкает в углу лаборатории. Здесь, торцом приставленный к подоконнику, стоит плоский стеклянный улей, в котором живет небольшая семья пчел. Ее поселяют сюда весной, перед тем как зацветают сады, и пчелы скоро
Природа власти
Природа власти ЧЕЛОВЕКООБРАЗНЫЕ ОБЕЗЬЯНЫ Их группы численно невелики и построены довольно просто, но по-разному у разных видов — от семейной у живущих на деревьях орангутанов до небольшого стада у шимпанзе, ведущих полуназемный образ жизни. Зоологи потратили много сил
8 ПРИРОДА ЧЕЛОВЕКА
8 ПРИРОДА ЧЕЛОВЕКА Вы хотите жить "согласно с природой"? О благородные стоики, какой обман слов! Вообразите себе существо, подобное природе, — безмерно расточительное, безмерно равнодушное, без намерений и оглядок, без жалости и справедливости, плодовитое и бесплодное, и
Природа света
Природа света 1. Между физиками уже давно существует разногласие о природе света. Одни полагают, что свет выбрасывается светящимися телами, тогда как другие думают, что он происходит от колебаний бесконечно тонкой упругой жидкости, распространенной во всем пространстве,
Как природа работает
Как природа работает А ведь это интересно — действительно, зачем безмозглый создал мозг?Вот я и задумался, есть ли еще какой-то смысл в существовании природы, преследует ли этот безмозглый какую-то свою цель между было и не было?«Цель безмозглого» — звучит глупо? Да,