Глава 7 Генетическая регуляция развития
Глава 7
Генетическая регуляция развития
У природы столько дел в этом мире, ей приходится создавать такую массу разнообразнейших творений, что по временам она и сама не в силах разобраться во всех тех различных процессах, которыми она одновременно занимается.
Уилки Коллинз «Женщина в белом»
Мутации и изменения, происходящие в онтогенезе
Если морфология представляет собой проявление сложного комплекса процессов развития, то эти процессы в свою очередь представляют собой проявление действия целого созвездия генов. Таково важное допущение, положенное в основу этой книги, и мы собрали по крайней мере prima facie данные в пользу того, что у Metazoa часть генома специфически участвует в регуляции онтогенеза и что характер эволюции этой части отличается от характера эволюции структурных генов. До сих пор, однако, мы ограничивались изолированными частными примерами генного контроля морфогенеза и не пытались ответить на центральный вопрос: каким образом гены определяют процесс развития?
Здесь мы в весьма прямом смысле возвращаемся к программе механики развития, выдвинутой Ру (Roux), однако, вместо того чтобы удалять клетки или другие структуры зародыша, с тем чтобы определить их роли в развитии, как это делали представители классической экспериментальной эмбриологии, генетика развития использует мутации как очень тонкий скальпель, дающий возможность уничтожать или изменять отдельные гены.
Генетическая модель, с помощью которой можно изучать любую систему, теоретически находящуюся под генетическим контролем, состоит в следующем. Для того чтобы проанализировать какой-либо процесс, в данном случае онтогенез, исследователь выявляет мутации, изменяющие этот процесс. Выявив такие мутации, он проводит фенотипическое сравнение мутантных особей с нормальными. Это сравнение помогает ему понять, как данный ген влияет на нормальное развитие. Однако, прежде чем продолжить описание метода проведения такого сравнения, следует указать, что воздействие мутаций на онтогенез проявляется двумя основными способами. Это, во-первых, дизруптивные изменения, при которых процесс нормального развития нарушается, что приводит к морфологическим аномалиям (например, к отсутствию некоторых структур). В наиболее резко выраженной форме такие мутации оказываются летальными. Во-вторых, это гомеозисные изменения, при которых под действием мутации развитие отклоняется от нормы, в результате чего какая-либо структура данного организма замещается гомологичным органом или конечностью. Мы отложим дальнейшее обсуждение мутаций этого второго типа до следующей главы и займемся здесь главным образом дизруптивными изменениями.
Анализ нарушений, вызываемых какой-либо дизруптивной мутацией, лишь в редких случаях сводится к простому сравнению конечного фенотипа гибнущей особи с нормальным фенотипом, потому что развитие - это сложный и высокоинтегрированный процесс. Огромное большинство происходящих в нем событий тесно связано с другими событиями и, в сущности, зависит от них. Это особенно ясно проявляется в том, что многие мутации обладают плейотропным действием, т. е. отсутствие или изменение одного гена может обусловить несколько морфологических изменений. Примером служат изменения, наблюдаемые у людей с так называемой аномалией Пельгера (Pg). Она наследуется у человека как простой доминантный аутосомный признак. У гетерозигот (Pg/+) нет никаких клинических симптомов, но для их нейтрофилов характерны аномально сегментированные ядра (рис. 7-1). У взрослого человека ядра полиморфноядерных нейтрофилов обычно состоят из четырех или пяти сегментов, у гетерозигот же Pg/+ ядра состоят всего из двух, реже из трех сегментов. Этот же признак обнаружен у кроликов, у которых он наследуется по тому же типу и сходным образом проявляется в картине крови. Скрещивая гетерозиготных кроликов, можно получить гомозиготных особей Pg/Pg. Ядра нейтрофилов у этих особей вообще не разделены на сегменты, и такой генотип сопровождается низкой жизнеспособностью. Для немногих выживших особей помимо этой особенности нейтрофилов характерна крайне выраженная карликовость с недоразвитием конечностей и грудной клетки (рис. 7-1). Здесь следует задать вопрос: какова причинная зависимость, если она существует, между этими двумя плейотропными нарушениями? Не исключена возможность, что оба этих фенотипических проявления представляют собой на самом деле результат третьего, пока еще неизвестного нарушения, вызванного аллелем Pg.
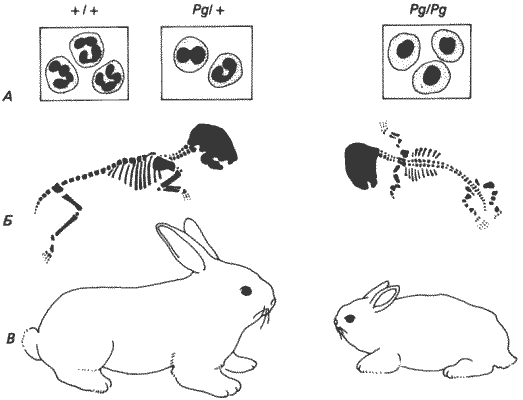
Рис. 7-1. Аномалия Пельгера у кроликов. А. Полиморфноядерные нейтрофилы нормальных ( +/+ ), гетерозиготных (Pg/+) и гомозиготных мутантных (Pg/Pg) кроликов. Б и В. Скелеты и общий вид взрослых кроликов Pg/+ (слева) и Pg/Pg(справа). Обратите внимание на резко выраженную недоразвитость конечностей у гомозиготных особей (Nachtsheim, 1950).
Широкий диапазон плейотропных воздействий наблюдается и при другом наследуемом изменении крови - при серповидноклеточной анемии. Она также наследуется у человека как простой аутосомный доминантный признак, отличаясь от аномалии Пельгера только тем, что нам точно известно вызывающее ее биохимическое нарушение. У людей, страдающих этим заболеванием, в ?-цепях гемоглобина замещена аминокислота в положении 6. В условиях низкого напряжения кислорода эта единственная замена изменяет конформационные свойства образующегося гемоглобинового тетрамера. В капиллярах и в мелких венах эритроциты, содержащие такой мутантный гемоглобин, деформируются, приобретая характерную «серповидную» форму. Такое изменение формы влечет за собой два непосредственных следствия. Во-первых, организм распознает аномальные серповидные эритроциты и разрушает их, что приводит к развитию анемии. Во-вторых, серповидные клетки часто закупоривают капилляры, нарушая кровоснабжение в отдельных участках, а тем самым нормальный рост и функционирование соответствующих органов.
Многочисленные и разнообразные нарушения, вызываемые этой заменой одной аминокислоты, схематически представлены на рис. 7-2. Глядя на этот рисунок, нетрудно понять, что неосведомленный человек, рассматривая схему снизу, может истолковать сам синдром совершенно иначе, чем если бы ему была известна первопричина всех этих нарушений.
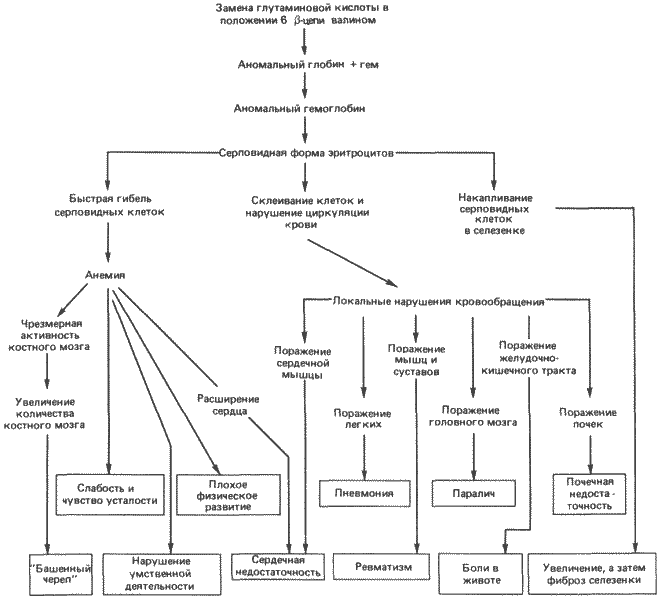
Рис. 7-2. Плейотропные эффекты замены одной аминокислоты в ?-цепи гемоглобина Homo sapiens, которая приводит к развитию серповидноклеточной анемии (Strickberger, 1976).
Сталкиваясь со сложным фенотипом, образовавшимся к концу ветвящегося и переплетающегося своими ветвями онтогенетического пути, необходимо помнить о существовании дихотомии. Геном можно теоретически разделить на две части. Одна его часть состоит из тех генов, которые определяют так называемые жизненно важные (housekeeping) функции, а другая - из генов, непосредственно участвующих в детерминации, дифференцировке и морфогенезе. Жизненно важные функции - это те обычные метаболические процессы и процессы поддержания жизнедеятельности клетки, которые, хотя и не участвуют прямо в морфогенезе, создают биохимические условия, абсолютно необходимые для жизни. Мутантный индивидуум, у которого отсутствует одна из транспортных РНК (тРНК) или ДНК-полимераза, встречается с серьезными морфогенетическими проблемами. Однако такого рода нарушения основного метаболизма не обязательно затрагивают какой-нибудь конкретный орган, ткань или стадию развития. Такие мутанты следует отличать от тех, которые оказывают прямое воздействие на развитие.
Прежде чем можно будет прийти к каким-то определенным выводам о природе генетических дефектов, необходимо затронуть еще два очень важных момента. Первый из них касается первичного места действия данного гена. Иными словами, существует ли некая специфичная ткань или орган, в которых ген проявляет свою активность? Более того, автономен ли ген в своей активности? Этот вопрос связан с тем, что на самом деле существует плейотропия двух типов: 1) в случае относительной, или зависимой, плейотропии, как при описанной здесь серповидноклеточной анемии, существует одно первичное место действия гена (т. е. эритроциты), а все остальные наблюдаемые нарушения связаны с этим одним дефектом или возникают как его следствие; 2) в случае прямой, или истинной, плейотропии все разнообразные дефекты, возникающие в различных тканях и/или органах, вызываются непосредственным действием одного гена. Грюнберг (Grunberg) на основании своих исследований мутаций у мышей считал преобладающим типом относительную плейотропию. Однако известны также примеры прямых плейотропных эффектов.
Второй момент, который необходимо учитывать при любом серьезном обсуждении генетической регуляции развития, - это время, когда данный ген осуществляет свою активность. Когда начинается эта активность, продолжается ли она непрерывно, занимает ли она один дискретный интервал времени или несколько таких интервалов? Для ответа на этот сложный вопрос необходимо, конечно, установить характер продукта изучаемого гена.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава XIII Регуляция экспрессии генов
Глава XIII Регуляция экспрессии генов О проблеме регуляции экспрессии генов мы в этой книге говорим фактически во всех главах, рассматривая ее с разных сторон. Существует такое, может быть несколько одностороннее, определение развития: «Понять развитие — это значит
Глава 6. Два энергетических принципа биологического развития
Глава 6. Два энергетических принципа биологического развития В особой земной оболочке, которой является биосфера, характеризуемая необратимыми процессами, жизнь будет увеличивать, а не уменьшать с ходом времени свободную энергию этой оболочки. В. И. Вернадский 6.1.
Глава 11. Генетическая эволюция продуктов поведения животных
Глава 11. Генетическая эволюция продуктов поведения животных Что именно мы понимаем под фенотипическим эффектом гена? Поверхностное знание молекулярной биологии может предложить нам такой ответ: каждый ген кодирует синтез одной полипептидной цепи, и в простейшем смысле
Глава 1. Генетическая информация
Глава 1. Генетическая информация Ключевой проблемой биологии, по-видимому, можно считать вопрос о том, как увековечивает свой опыт живая материя. М. Дельбрюк (1906–1981), американский генетик, лауреат Нобелевской премии 1969 г. При изучении любого биологического феномена
11.5. Генетическая психофизиология
11.5. Генетическая психофизиология Генетическая психофизиология изучает взаимодействие генотипа и факторов среды в формировании физиологических систем организма, обеспечивающих психическую деятельность (Булаева К. Б., 1991).Структурная единица нервной системы – нейрон –
Глава 13. Генетика развития[1]
Глава 13. Генетика развития[1] Пусть не покажется странным, если я позволю себе сказать, что легче понять образование всех небесных тел и причину их движений, короче говоря, происхождение всего современного устройства мироздания, чем точно выяснить на основании механики
Глава 4. Регуляция активности ферментов. Медицинская энзимология
Глава 4. Регуляция активности ферментов. Медицинская энзимология Способы регуляции активности ферментов:1. Изменение количества ферментов.2. Изменение каталитической эффективности фермента.3. Изменение условий протекания реакции.Регуляция количества
Глава 27. Регуляция и взаимосвязь метаболизма
Глава 27. Регуляция и взаимосвязь метаболизма Для нормального функционирования организма должна осуществляться точная регуляция потока метаболитов по анаболическим и катаболическим путям. Все сопутствующие химические процессы должны протекать со скоростями,
ГЛАВА 1 ПЕРИОДЫ РАЗВИТИЯ ЩЕНКОВ
ГЛАВА 1 ПЕРИОДЫ РАЗВИТИЯ ЩЕНКОВ Роды завершают эмбриональный период развития щенка. Послеутробный период развития животных начинается с их рождения и кончается естественной смертью в результате старения.Каждый период развития, состоящий из ряда определенных фаз и
Глава 3. Генетическая информация
Глава 3. Генетическая информация Носителями наследственной информации в природе являются нуклеиновые кислоты. Именно они выполняют три необходимые для жизни функции: хранение, воспроизведение и реализацию этой информации. В ходе эволюции ключевая роль по хранению и
Глава 4. Среда развития
Глава 4. Среда развития Сторожевые пастушьи собаки «К западу от Маньчжурии вплоть до Венгерской равнины простирался океан травы, по его холмистым пространствам кочевники перегоняли свои стада в постоянном поиске пищи. Зимой они укрывались от буранов и метелей в горах,
Глава 10 Адаптации экспрессии генов в процессе развития
Глава 10 Адаптации экспрессии генов в процессе развития Жизнь -это сила, которая проделывает бесчисленное множество экспериментов, пытаясь организовать себя ... мамонт и человек, мышь и мегатерий, мухи и отцы церкви - все это результаты более или менее успешных попыток
Генетическая стоимость развития на молекулярном уровне
Генетическая стоимость развития на молекулярном уровне Попытки перенести оценки числа генов, необходимых для развития дрозофилы, на другие организмы сталкиваются с двумя проблемами. Во-первых, геном дрозофилы, по крайней мере в некоторых существенных отношениях,
5. Теория стадийного развития и особенности развития животных
5. Теория стадийного развития и особенности развития животных В основе управления развитием организмов лежит теория стадийного развития, которую сформулировал академик Т. Д. Лысенко, исходя из работы И. В. Мичурина и многочисленных собственных исследований.Несмотря на
Глава 5. Проблемы индивидуального развития и пол
Глава 5. Проблемы индивидуального развития и пол Генетические аномалии полового развития и половая идентификация В рамках онтогенетической концепции полаДж. Мани полноценный мужчина или женщина должны в своем индивидуальном развитии демонстрировать все восемь