Разрывы, недостающие звенья и эволюционные механизмы
Разрывы, недостающие звенья и эволюционные механизмы
Отсутствие предковых форм или форм, промежуточных между отдельными ископаемыми видами, не является некой странной особенностью ранней истории Metazoa. Разрывы представляют собой общее явление и встречаются по всей палеонтологической летописи. Дарвина смущало отсутствие в палеонтологической летописи непрерывного ряда промежуточных в эволюционном отношении форм, поскольку, согласно его теории, наличия таких переходных форм следовало ожидать: «...в таком случае число существовавших когда-то промежуточных разновидностей должно быть поистине огромным. Почему же тогда каждая геологическая формация и каждый слой не переполнены такими промежуточными звеньями?» В «Происхождении видов» Дарвин затратил много усилий, пытаясь ответить на этот вопрос и дать ему рациональное объяснение. Он выдвинул три основные причины. Первая из них - неполнота палеонтологической летописи. Разрывы вызваны полным уничтожением ископаемых остатков в результате эрозии и других процессов или же несохранением когда-то существовавших промежуточных форм.
Вторая причина состоит в искажении летописи таксономическими артефактами, создаваемыми самими исследователями. Интерградирующим формам иногда присваивают видовые названия, затемняя таким образом их переходный характер. Современные статистические и стратиграфические методы позволяют избегать таксономических артефактов. Здесь уместно, однако, привести один важный пример. До недавнего времени покрытосеменные из меловых отложений относили к современным родам на основании формы листьев; в результате возникновение современных родов цветковых растений представлялось более внезапным, чем это было на самом деле, о чем свидетельствуют новые исследования морфологии ископаемых листьев и цветков, как это обсуждается в работах Дильхера (Dilcher).
Третья причина, по Дарвину, заключается в том, что разрывы могут быть следствием природы самого эволюционного процесса. Эволюционные превращения, согласно Дарвину, вероятно, обычно происходили в небольших географически ограниченных популяциях, после чего новая форма быстро распространялась по более обширному ареалу предкового вида, или «...период, в течение которого каждый вид претерпевал модификации, хотя он длителен, если измерять его в годах, был, возможно, невелик по сравнению с тем периодом, в течение которого он не претерпевал никаких изменений». Локальные или быстро эволюционирующие популяции лишь в редких случаях могут сохраниться в палеонтологической летописи.
Палеонтологическая летопись очень необъективна. В некоторых обстановках, таких как мелководные морские бассейны, образование отложений, содержащих ископаемые остатки, более вероятно, чем в других, таких как горные хребты. Некоторые организмы сохраняются лучше, чем другие. Например, моллюски, позвоночные и иглокожие очень хорошо представлены в палеонтологической летописи, насекомые представлены в ней довольно слабо, а от планарий и нематод, чрезвычайно широко распространенных в современных фаунах, практически не осталось следов. Однако разрывы существуют даже в палеонтологической летописи тех типов, история которых отражена в ней очень хорошо.
В 1959 г., в статье, посвященной столетию дарвиновского «Происхождения видов», Симпсон (G. G. Simpson) писал, что, несмотря на продолжавшееся все это время интенсивное изучение ископаемых форм и многочисленные их находки, известная нам часть палентологическои летописи дает лишь очень слабое и неадекватное представление о существовавшей в прошлом жизни. Симпсон привел результаты очень интересного «бумажного эксперимента», который он проделал, чтобы выяснить возможные последствия неполноты летописи. Эксперимент состоит в следующем: берут случайную выборку, содержащую 10% видов, относящихся к гипотетическому филогенетическому древу, которое состоит из нескольких семейств, делящихся на ряд родов, подразделяющихся в свою очередь на многочисленные виды. Виды, включенные в выборку, отображают неполноту палеонтологической летописи. Эти «найденные» виды распределяют по родам и семействам, как если бы они были настоящими ископаемыми остатками. Как и можно было ожидать для небольшой случайной выборки, большая часть попавших в нее видов никак не была связана друг с другом, так что лишь в нескольких случаях удалось построить непрерывный ряд видов, представляющих собой прямую последовательность от предков к потомкам. Средняя длина разрывов (число отсутствующих «неизвестных» видов между «найденными» видами) возрастает с повышением таксономического ранга, так что между всеми обнаруженными семействами получаются разрывы, соответствующие большому числу видов. Получилась картина, сходная с настоящей палеонтологической летописью, где разрывы между группами высших таксономических рангов наблюдаются всегда и обычно бывают велики.
Ввиду близкого соответствия характера разрывов, обнаруженных в этом эксперименте, и разрывов, действительно наблюдаемых в палеонтологической летописи, Симпсон пришел к заключению, что палеонтологическая летопись представляет собой небольшую случайную выборку из первоначально непрерывных филогенетических последовательностей. Исходя из этого постулата, Симпсон сделал ряд предсказаний.
1. Ископаемые остатки, относящиеся к крупному разрыву, должны обнаруживаться лишь в очень редких случаях. Наилучшим примером такого рода служит археоптерикс, так хорошо заполнивший разрыв между рептилиями и птицами.
2. Ряды последовательных родов должны встречаться чаще, чем ряды последовательных видов. Это объясняется тем, что в выборки попадают преимущественно представители крупных широко распространенных видов. Однако в отличие от «бумажного эксперимента», в котором вероятность попасть в выборку была одинакова для всех видов, в реальных родах частота мелких локализованных видов может оказаться выше. Но, хотя виды, образующие многочисленную популяцию, имеют больше шансов сохраниться в палеонтологической летописи, вероятность дать начало новому роду для них не выше, чем для видов с малочисленной популяцией. В таких случаях нам будут известны последовательные роды, но лишь по видам, которые сами не укладываются в прямую последовательность.
3. Некоторые крупные таксоны будут вновь обнаруживаться после долгого отсутствия. Наилучшим известным примером этого служит одна из ветвей кистеперых рыб - Coelacanthidae. Последние ископаемые целаканты относятся к меловому периоду, однако вблизи побережья Мадагаскара до сих пор процветают ныне живущие представители этой группы. Другие таинственные долгожители - это Neopilina - единственный современный представитель моллюсков класса Monoplacophora, последний раз обнаруженного в палеонтологической летописи в силурийских отложениях, и Platasterias - представитель подкласса Somasteroidea (иглокожие), последний раз найденного среди девонских ископаемых остатков. Появление этих классов после их отсутствия на протяжении 300 или 400 млн. лет свидетельствует о степени несовершенства палеонтологической летописи и, по-видимому, подтверждает справедливость точки зрения Симпсона о том, что разрывы могут быть результатом случайностей, связанных со сбором материала и его сохранностью.
В 1972 г. Элдридж и Гулд (Eldredge, Gould) высказали мнение, что нельзя относить за счет неполноты все разрывы палеонтологической летописи, потому что даже у тех видов, которые встречаются в ней в течение длительных периодов времени, обычно не наблюдается постепенных эволюционных изменений в одном направлении и они остаются, по существу, неизмененными на протяжении всей своей истории. Виды-потомки часто появляются внезапно, создавая ясно выраженную прерывистость. Эти авторы полагают: «Если новые виды возникают очень быстро как небольшие периферически изолированные локальные популяции, то в таком случае надежды обнаружить последовательные ряды незаметно переходящих друг в друга ископаемых остатков несбыточны. Новый вид развивается не в той области, где обитали его предки; он возникает не в результате медленного превращения всех своих предшественников. Многие разрывы палеонтологической летописи отражают то, что имело место в действительности». Таким образом, «большинство эволюционных изменений в морфологии происходит в течение времени, короткого по сравнению с общей продолжительностью существования видов». Если для Дарвина это было лишь одной из ряда возможных причин существования разрывов, то Элдридж и Гулд считали это главной его причиной. Процесс эволюции осуществляется не путем постепенного изменения всей видовой популяции. Сначала происходит быстрое изменение небольшой периферической популяции, отделенной от основной. Это эволюционное изменение сопровождается видообразованием. В дальнейшем в какой-то период этот новый вид может распространиться и заместить основную популяцию предкового вида по всему его ареалу. Исследователю, изучающему палеонтологическую летопись, такая последовательность событий покажется внезапным разрывом: виды-потомки возникают без всяких признаков эволюционного перехода от своих предковых видов. Это не означает, что переходных форм не существовало, что эволюция совершается скачками; это лишь значит, что переходные популяции имели низкую численность, что они существовали недолго и что их географическое распространение было ограничено небольшой областью на окраине ареала основной предковой популяции. Поэтому у них было мало шансов сохраниться в виде ископаемых остатков.
Рассуждая таким образом, Элдридж и Гулд смогли связать разрывы, столь многочисленные в палеонтологической летописи, с современными концепциями, согласно которым аллопатрическое видообразование, затрагивающее мелкие локальные популяции, представляет собой обычный способ видообразования и может происходить очень быстро. Скорости и способы видообразования будут рассмотрены в гл. 3 после более исчерпывающего рассмотрения всего спектра типов эволюционных скоростей и их механистических взаимоотношений.
Хотя модель Элдриджа и Гулда теоретически приемлема и согласуется с существованием разрывов, наличие в палеонтологической летописи разрывов или стабильных видов в промежутках между разрывами никоим образом не доказывает ее правильности. Прерывистая эволюция обладает известным сходством с детективным романом. Все интересные события скрыты от глаз, они происходят «за сценой», в ускользающих от наблюдения периферических популяциях. Необходимы реальные примеры прерывистой эволюции. Это тем более необходимо, что если даже модель Элдриджа и Гулда верна и соответствует преобладающему или даже обычному способу эволюции, то палеонтологическая летопись тем не менее остается несовершенной и последствия этого несовершенства, на которые указал Симпсон, сохраняют силу. Вероятный пример прерывистой эволюции описала Овчаренко (1969), изучавшая эволюцию двух обычных и широко распространенных видов юрских брахиопод - Kutchithyris acutiplicata и происходящего от него К. euryptycha. Стратиграфически К. euryptycha встречается выше К. acutiplicata. В одной ограниченной местности Овчаренко нашла пласт толщиной 1,0-1,5 м, в нижней части которого содержались только К. acutiplicata, а в верхней-только К. euryptycha. Однако между этими двумя частями была тонкая (толщиной 10 см) прослойка, в которой присутствовали оба вида, а также промежуточные формы. В статье, опубликованной в 1972 г., Элдридж и Гулд привели два примера из своих собственных исследований, подтверждающие их гипотезу. Эти примеры интересны не только потому, что они относятся к реальным линиям (последовательным рядам) ископаемых, интерпретируемых с точки зрения прерывистой эволюции, но также потому, что они позволяют обсудить возможные эмбриогенетические основы наблюдаемых эволюционных изменений.
Первый пример, изучавшийся Гулдом, это эволюция Poecilozonites bermudensis zonatus - наземной улитки, относящейся к последним 300 000 лет плейстоцена Бермудских островов. Ископаемый подвид превосходно сохранился, а кроме того, имеется ныне живущий подвид, с которым его можно сравнивать. Две длительно существовавшие восточная и западная популяции P. b. zonatus дали начало педоморфным ветвям. Педоморфозом называют сохранение у половозрелых животных признаков, характерных для ювенильных стадий их предковых форм. Раковины взрослой стадии ггедоморфного подвида сходны с раковинами юной стадии предкового подвида по типу окраски, общей форме спирали, толщине раковины и форме губы устья. На основе подробного изучения географических, стратиграфических и морфологических характеристик педоморфных подвидов Гулд считает, что они не относятся к одной непрерывной педоморфной линии. По его мнению, дело обстояло иначе: исходные популяции P. b. zonatus дали начало нескольким последовательным педоморфным линиям в ответ на периодически повторяющиеся изменения условий среды, приведшие к образованию почв, бедных известью, которые благоприятствовали тонким раковинам ввиду низкого содержания в них кальция. Такие линии возникали быстро, а наиболее доступным путем к тонким раковинам был педоморфоз.
Если Гулд правильно интерпретировал эти данные, то интересно отметить, что генетические изменения, необходимые для такой модификации развития какого-либо организма, которая бы привела к педоморфозу, по-видимому, минимальны. В наиболее хорошо изученном примере педоморфоза - у хвостатых амфибий рода Ambystoma - генетической основой для определения того, пойдет ли развитие данной особи по пути метаморфоза или педоморфоза, служит пара аллелей одного гена (см. гл. 6). Если допустить, что аналогичная регуляция имеет место в развитии брюхоногих моллюсков, то первоначальное генетическое изменение, направившее их эволюцию по пути педоморфоза, могло произойти в небольшой популяции очень быстро.
Вторым примером, представленным Элдриджем и Гулдом, была эволюция подвидов среднедевонскою трилобита Phacops rana (рис. 2-3), которую изучал Элдридж. Очень крупные глаза Phacops состоят из многочисленных крупных дискретных фасеток, расположенных рядами, тянущимися в дорсовентральном направлении. Главные эволюционные изменения, наблюдаемые у подвидов Р. rana, - это уменьшение числа таких дорсовентральных рядов от 18 у самого древнего подвида до 17, а в дальнейшем - до 15 у более молодых подвидов. В пределах отдельных подвидов число рядов обычно оставалось стабильным, хотя обнаружены популяции, у которых оно варьировало. Различия в числе рядов между разными подвидами носят характер внезапных разрывов, где прежнее число рядов сменяется новым стабильным их числом. Элдридж считает, что эти события происходили аллопатрически в периферических популяциях. Последующее распространение нового подвида с числом рядов, стабилизировавшимся на новом уровне, было внезапным, если иметь в виду характер его отражения в палеонтологической летописи.
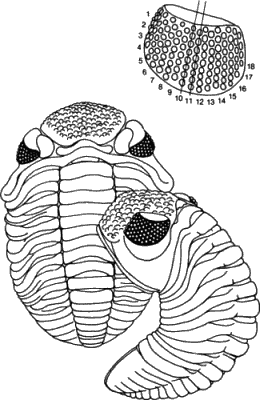
Рис. 2-3. Девонский трилобит Phacops и организация фасеток в его глазу. У этого подвида, P. rana crassituberculata, имелось 18 дорсовентральных рядов фасеток (Levi-Setti, 1975).
При такой интерпретации не возникает никаких генетических проблем, потому что, хотя можно было бы думать, что генная регуляция такого сложного признака, как число рядов фасеток в глазу, должна быть также сложной, это на самом деле не так. Непосредственное изучение генетики трилобитов, к сожалению, исключено, но исследование регуляции таких меристических признаков, как число щетинок у дрозофилы или число пальцев у млекопитающих, показывает, что в их регуляции участвует лишь небольшое число генов (локусов) (см. гл. 5). Так, например, замещение нескольких аллелей у морских свинок вызывает у них скачкообразные изменения числа пальцев. Если число рядов фасеток у Phacops регулировалось аналогичным образом, то замещение аллелей, а тем самым изменение числа рядов фасеток могло произойти в результате эффекта основателя, возникшего в небольшой популяции. Необходимое для этого генетическое изменение очень невелико и (в геологическом масштабе времени) могло произойти мгновенно. Скромные размеры генетического изменения, которое, вероятно, потребовалось для того, чтобы произошли морфологические преобразования, отмеченные у Poecilozonites и Phacops, не означают, что подобные события незначительны. Напротив, «легкие» генные изменения могут послужить быстрым начальным стимулом для дальнейших генетических изменений в эволюционирующей популяции.
В 1977 г. Гулд и Элдридж пересмотрели гипотезу прерывистой эволюции в свете результатов палеонтологических исследований, проведенных после того, как в 1972 г. была опубликована созданная ими модель. Они смогли привести ряд случаев, относящихся к радиоляриям, аммонитам, трилобитам и даже гоминидам, для которых объяснение на основе прерывистой эволюции представляется наиболее вероятным. По их мнению, лишь одно исследование - работа Озавы по эволюции одной фораминиферы из перми - представляет собой несомненный пример постепенной эволюции целой популяции. Букштейн (Bookstein), Гингерих (Gingerich) и Клюге (Kluge) горячо оспаривали некоторые из примеров, приводимых Гулдом и Элдриджем: проделанный ими повторный анализ данных Гингериха об эволюции некоторых эоценовых млекопитающих привел их к выводу, что в 12 случаях эти данные свидетельствуют о постепенном изменении, в 4х - о прерывистости и в одном - о застое. Подобным же образом Кронин (Cronin) недавно проанализировал имеющиеся данные об эволюции гоминид и пришел к заключению, что эволюцию человека можно с большим основанием интерпретировать как постепенную, с периодами, когда развитие ускорялось, и периодами, когда оно замедлялось. Концепция прерывистости, возможно, остается пригодной для объяснения исходной дивергенции гоминид от человекообразных обезьян. Различия между человеком и шимпанзе, обнаруженные на молекулярном уровне, свидетельствуют в пользу быстрой дивергенции, однако в настоящее время соответственные палеонтологические данные отсутствуют.
Для того чтобы с уверенностью утверждать, что эволюция данной линии организмов была прерывистой, необходимо располагать непрерывными палеонтологическими данными за длительное время, которые позволили бы распознать как периоды застоя, так и периоды быстрой эволюции, причем необходимо иметь возможность точно датировать (по абсолютной шкале) короткие интервалы в пределах этой последовательности. Эти условия были соблюдены в недавнем подробном исследовании эволюционной истории кайнозойских пресноводных моллюсков из Турканского бассейна (Восточная Африка), проведенного Уильямсоном (P. G. Williamson). Уильямсон изучил тысячи ископаемых остатков, относящихся к 13 линиям брюхоногих и двустворчатых моллюсков из мощной толщи осадочных пород, прослоенных вулканическими туфами точно установленного возраста. Многие из этих видов оставались неизменными на протяжении нескольких миллионов лет, и ни в одной линии не наблюдалось постепенных морфологических изменений. Вместо этого в периоды стрессовых ситуаций, возникавших в результате усыхания озера, в котором они обитали, относительно быстро появлялись новые виды. В периоды таких стрессов эти моллюски были, возможно, изолированы от других представителей своего вида, населявших другие озера. Стратиграфические данные Уильямсона были достаточно точны, а его коллекции ископаемых экземпляров достаточно велики, так что он имел возможность документировать переходные моменты в тех линиях, которые изучал. Он обнаружил, что в промежуточных популяциях изменчивость морфологических признаков была выше, чем в уже сложившихся видах. По его мнению, эта изменчивость вызвана частичным разрушением морфогенетического гомеостаза, что ведет к большему фенотипическому разнообразию и быстрой морфологической эволюции.
Конечно, не возникает сомнений в том, что у моллюсков из Турканского бассейна налицо все признаки прерывистой эволюции - длительные периоды застоя с эпизодическими всплесками относительно быстрого развития. А с какой именно скоростью протекает относительно быстрая эволюция? Согласно Уильямсону, эпизодические всплески занимали от 5 до 50 тыс. лет. Учитывая точность, которая обычно достигается при изучении палеонтологической летописи, этот промежуток времени и в самом деле невелик. Как отметил Джонс (J. S. Jones), обсуждая наблюдения Уильямсона с точки зрения генетики, для самих организмов 5-50 тыс. лет - время немалое. У ныне живущих родичей этих моллюсков время генерации колеблется от шести месяцев до года, т. е. для наблюдавшихся морфологических изменений в среднем потребовалось 20000 поколений. Как указывает Джонс, это эквивалентно тысячелетнему эксперименту на дрозофиле, или селекционному эксперименту на мышах, продолжительностью в 6000 лет, или выведению в течение 40000 лет породы собак или других домашних животных. В обычных селекционных экспериментах резкие морфологические изменения и даже репродуктивная изоляция возникают иногда всего за 20-50 поколений. Моллюски, которых изучал Уильямсон, развивались не особенно быстро, но они хорошо иллюстрируют концепцию прерывистого равновесия.
Полемика, возникшая среди палеонтологов в связи с этой концепцией, занимательна, однако сама гипотеза прерывистости представляется правильной, по крайней мере в некоторых случаях. Как показал Харпер (Harper), прерывистая эволюция и филетический градуализм - это два крайних случая целого спектра возможностей; по-видимому, имели место и тот, и другой. Суть дела, конечно, не в том, исключают ли прерывистость и градуализм друг друга, а в том, приводит ли один из этих способов к количественно более значительной или качественно иной морфологической эволюции, чем другой, и что означают эти выявленные палеобиологами способы эволюции, если рассматривать их в аспекте эмбриогенетических процессов, лежащих в основе морфологической эволюции.
Аммониты - вымершие головоногие, раковины которых были разделены на камеры, подобно раковинам ныне живущих наутилусов, - служат примерами, подтверждающими обе гипотезы. Последовательный ряд аммонитов, показанный на рис. 2-4, эволюционировал на протяжении примерно 3 · 106 лет, и в нем выявляется несколько тенденций: постепенное увеличение, а затем постепенное уменьшение размеров; постепенное свертывание раковины, в результате чего жилая камера постепенно все плотнее прилегает к остальной части раковины; усложнение характера лопастной линии, сменяющееся уменьшением ее сложности и некоторыми качественными изменениями ее формы; и наконец, постепенное усиление, а затем ослабление ребристости. Эти тенденции привели лишь к мелким изменениям как в ту, так и в другую сторону, а многие из них даже ревертировали на протяжении времени существования этого ряда форм.
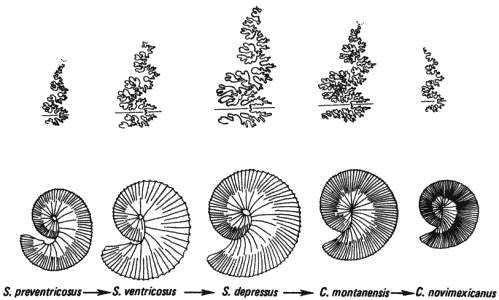
Рис. 2-4. Эволюционный ряд аммонитов, демонстрирующий изменения размеров раковины, ее формы, ребристости и характера лопастной линии, происходившие на протяжении примерно 3 млн. лет. Над каждой раковиной изображена форма лопастной линии, которая определяет прикрепление стенок газовых камер к внутренней поверхности раковины. Линия позднемеловых аммонитов тянется от самого древнего вида Scaphites preventricosus через S. ventricosus, S. depressus к Clioscaphites montanensis и С. novimexicanus. (Cobban, 1951.)
Совершенно иной набор эволюционных событий изображен на рис. 2-5, где представлено предполагаемое эволюционное происхождение гетероморфных аммонитов от предков с раковиной, закрученной обычным образом, за короткий промежуток времени в позднем триасе. Длительность всего норийского яруса равна примерно 5-10 · 106 лет. Глубокие изменения, показанные на рис. 2-5, занимали лишь ограниченный промежуток времени и происходили гораздо быстрее, чем мелкие постепенные изменения в ряду, показанном на рис. 2-4. Другие группы юрских и меловых аммонитов также дали начало гетероморфам. Эволюция гетероморф с их принципиально различающимися типами закручивания раковины требовала значительной модификации морфогенетических процессов, а между тем создается впечатление, что они были достигнуты довольно быстро.
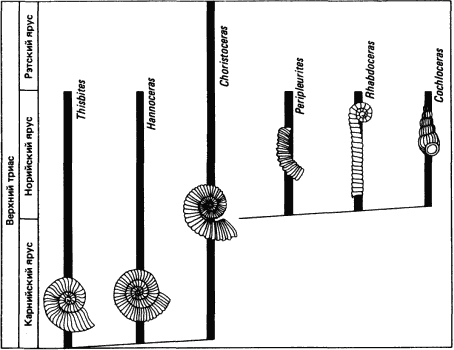
Рис. 2-5. Быстрая эволюция гетероморфных аммонитов от предков с нормальной раковиной в течение позднего триаса. Длина столбиков соответствует временным границам для каждого рода. Аммониты, которые были свободно плавающими животными, изображены в соответствии с ориентацией их тела при жизни. Типичные аммониты были платиспиральными. Эволюция гетероморфов сопровождалась изменениями характера роста, которые были необходимы для возникновения незакрученных раковин, как у Rhabdoceras, или раковин со спиральными завитками, как у Cochloceras (Weidemann, 1969).
Скорости эволюции можно изучать безотносительно к определенным ее формам (филетическая трансформация всей популяции или же видообразование в результате отделения мелких периферических популяций). Такой подход может оказаться очень информативным, как это становится очевидным при рассмотрении различных скоростей эволюции, сделанном Симпсоном (Simpson) в его книге «Главные черты эволюции», написанной в 1953 г. Симпсон различает три крупные категории скоростей эволюции: горотелические, брадителические и тахителические. К горотелическим относятся скорости, более или менее близкие к средней для данной группы организмов. Горотелия может оказаться эквивалентной градуализму Элдриджа и Гулда. Однако это необязательно, поскольку горотелическая скорость может сложиться в результате усреднения прерывистых событий, происходивших на протяжении длительного времени.
Брадителическими называют скорости морфологической эволюции, близкие к нулю. Всем хорошо знакомо выражение «живое ископаемое» - так называют организм, у которого произошло очень мало явных морфологических изменений за многие миллионы лет, тогда как родственные ему формы претерпели существенную эволюцию. Например, среди ныне живущих приматов имеется несколько очень примитивных представителей полуобезьян (Prosimii), которые не слишком сильно отличаются от своих (и наших) эоценовых предков. Наряду с ними существуют также различные специализированные полуобезьяны; разнообразные формы низших обезьян, более или менее продвинувшихся в своем эволюционном развитии; человекообразные обезьяны; форма, наиболее сильно дивергировавшая от исходного типа, - человек. Ясно, что примитивные ныне живущие полуобезьяны эволюционировали гораздо медленнее как морфологически, так, вероятно, и в отношении поведения, чем та линия гоминид, нынешней гордостью и украшением которой служит Homo sapiens.
Скорости эволюции могут резко изменяться в пределах одной линии. Превосходный пример содержится в работе Уэстола (Westoll) о скорости эволюции двоякодышащих рыб. По данным Уэстола (из книги Симпсона), построены графики, представленные на рис. 2-6. Ископаемым двоякодышащим рыбам были даны оценки, отражающие их близость к ныне живущим двоякодышащим, в зависимости от степени «модернизации» их морфологических признаков. Оценка 100 дана современным признакам, а 0 - самым примитивным. Относительная скорость эволюции определяется как изменение оценки за миллион лет. Эволюция двоякодышащих протекала относительно быстро (для двоякодышащих) в первые 50 · 106 лет истории этой группы, а в дальнейшем шла крайне медленно - так медленно, по сравнению с другими рыбами, что мы можем считать двоякодышащих живыми ископаемыми.
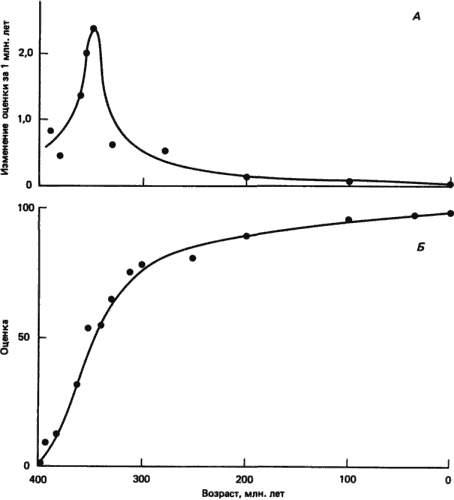
Рис. 2-6. Эволюция морфологических признаков у двоякодышащих. А. Скорость эволюции двоякодышащих. Б. Приобретение «современных» признаков (оценка для современных рыб 100 баллов). (Simpson, 1953; с изменениями.)
Брадителические скорости не означают отсутствия мутаций или отбора; они лишь означают, что отбор благоприятствует незначительному конечному морфологическому изменению или его отсутствию. Анализ степени белкового полиморфизма и генетической гетерозиготности был произведен для одного живого ископаемого - мечехвоста Limulus polyphemus (Selander et al., 1970). Ископаемые остатки подкласса Xiphosura, к которому принадлежит мечехвост, впервые обнаруживаются в среднем кембрии. Ископаемые мечехвосты различаются по местообитаниям (среди них были морские, солоноватоводные и пресноводные формы), что позволяет предполагать их физиологическую пластичность. Тем не менее морфологическая эволюция Xiphosura была достаточно консервативна. Позднепалеозойские, мезозойские и ныне живущие формы сходны по своим общим признакам. Сам Limulus плохо представлен в палеонтологической летописи, содержащей остатки третичных Xiphosura в исчезающе малых количествах. Однако ныне живущие мечехвосты очень сходны с юрским Mesolimulus, который, подобно Limulus, обитал в море; такое сходство свидетельствует об относительно медленной эволюции этой линии. Методом электрофореза был определен полиморфизм по 24 белкам, кодируемым 25 генетическими локусами. Оказалось, что доля полиморфных локусов в популяции мечехвоста сходна с их долей в популяциях мышей, дрозофилы и человека. У отдельных особей мечехвоста доля гетерозиготных локусов также оказалась сходной с их долей у отдельных особей тех видов, с которыми его сравнивали. Таким образом, брадителический мечехвост обладает такой же высокой генетической изменчивостью, как представители групп, эволюционирующих быстрее. Такая изменчивость обнаружена и у других брадителических организмов. В популяциях опоссумов, которые мало изменились после мелового периода, наблюдается такая же морфологическая изменчивость, как и у других млекопитающих. Кроме того, брадителические организмы обладают значительным запасом изменчивости, создающим возможность для видообразования, в некоторых случаях весьма обильного. Например, Selaginella - род растений, который, согласно Филипсу (Phillips) и Лейсману (Leisman), мало отличается по своему общему строению от пенсильванских плауновых (Lycopodiales) Paurodendron и Selaginellites, представлен в современной флоре 200 видами.
Симпсон (Simpson) считает, что брадителия поддерживается у организмов, занимающих адаптивную зону, которая сохраняется на протяжении исключительно долгого времени. Такая зона может быть узкой, и ее может занимать какой-либо хорошо приспособленный к ней организм, вроде двоякодышащих; или она может быть широкой, и тогда ее занимает такой всеядный организм, как опоссум. Существенные изменения, по-видимому, контролируются нормализующим (стабилизирующим) отбором, обеспечивающим сохранение высокой адаптированности. Иногда брадителические организмы дают начало быстро эволюционирующим формам. Это означает, что брадителические организмы обладают генетической пластичностью, благодаря которой они могут ответить на сильное давление отбора, вызывающего отклонения от нормы.
Самые высокие скорости эволюции относятся к третьей и, возможно, наиболее интересной категории - тахителическим скоростям. Симпсон утверждает: «Я считаю, что тахителия - обычный элемент, участвующий в возникновении высших категорий, и что она помогает объяснить пробелы, систематически встречающиеся в палеонтологической летописи». Тахителия сходна с прерывистостью в представлении Элдриджа и Гулда: в основе как той, так и другой концепции лежат исключительно высокие скорости эволюции. Однако Элдридж и Гулд делали упор на модель видообразования, исходящую из популяционной генетики, а Симпсон подходил к тахителии с другой, комплементарной, точки зрения. Он считал, что главное событие, сопутствующее тахителии, - это сдвиг популяции из одной крупной адаптивной зоны в другую.
Когда сдвигающаяся популяция переступает порог для перехода из одной зоны в другую, то это подразумевает, что на время перехода она оказывается в некой метастабильной зоне. В таких обстоятельствах она, вероятно, может выжить только при отсутствии конкуренции, с тем чтобы, будучи плохо приспособленной, она могла удержаться в новой зоне до тех пор, пока не достигнет удовлетворительного уровня адаптации. Таким образом, в ранний период радиации новых групп, распространяющихся в незанятые адаптивные зоны, возможна тахителия. Во время такой быстрой радиации все линии относительно плохо приспособлены и между ними нет конкуренции. В результате, как мы это видели на примере иглокожих, возникают разнообразные линии; одни из них быстро вымирают, тогда как другие линии укрепляют свои позиции в данной адаптивной зоне за счет своих менее удачливых родичей. Трудность достижения быстрых изменений в морфогенетических процессах, необходимых при тахителической эволюции, связана с тем, что, какими бы ни были экологические факторы, способствующие или не препятствующие переходу тахителической популяции из одной зоны в другую, какой бы степени ни достигала допустимая при этом рыхлость его адаптированности к окружающей среде, для того чтобы этот организм мог как-то существовать и размножаться, он должен продолжать оставаться морфогенетически и функционально интегрированным. Или, как это удачно выразил Фразетта (Frazzetta) в своих «Комплексных адаптациях у эволюционирующих популяций»: «Эволюционная проблема заключается, в сущности, в том, как постепенно совершенствовать машину, не прерывая ее работы».
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
6.3. Экспериментальные эволюционные машины (ЭЭМ)
6.3. Экспериментальные эволюционные машины (ЭЭМ) В предыдущем параграфе мы упоминали о том, что непрерывный рост микробных популяций в проточных системах позволяет экспериментально изучать микроэволюционные переходы, т.е. шаги эволюции.Суть непрерывного процесса
Глава 9 Что такое примат? Развитие со времен динозавров. Кометы и прочие эволюционные механизмы.
Глава 9 Что такое примат? Развитие со времен динозавров. Кометы и прочие эволюционные механизмы. Как я уже говорил, палеонтологический «пробел» измеряется четырьмя миллионами лет – целых сорок метров на нашей мерной ленте, где один миллиметр соответствует ста
3. ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА
3. ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА Строение мозга у животных разных видов разнообразно. И хотя, как следует из рис. 19.3, родственные виды, например среди ракообразных или млекопитающих, имеют общие черты в строении мозга, между собой они мало похожи. Именно с появлением
Разрывы мышц
Разрывы мышц При травматических разрывах мышц, как правило, животные страдают от интенсивных болей. Полные разрывы мыщц встречаются достаточно редко — в подобных случаях всегда требуется хирургическое вмешательство. Во всех случаях неполных разрывов (или надрывов)
Эволюционные истоки морфологических особенностей человека
Эволюционные истоки морфологических особенностей человека Как с эволюционной точки зрения интерпретировать переход к прямохождению, исчезновение волос на теле, нетипичное для обезьян распределение жировой ткани, стремительное увеличение размеров головного мозга и
Эволюционные стратегии популяций
Эволюционные стратегии популяций В эволюционной экологии рассматриваются различные теоретические обобщения, показывающие межвидовые взаимоотношения. В результате взаимодействия у животных выработались собственные эволюционные стратегии. Одной из важнейших
Эволюционные изменения в организации яиц со спиральным дроблением
Эволюционные изменения в организации яиц со спиральным дроблением Для многих зародышей со спиральным дроблением, в особенности для зародышей моллюсков и кольчецов, составлены очень точные карты, на которых указана дальнейшая судьба различных клеток. Разные типы
Эволюционные изменения в организации яиц хордовых
Эволюционные изменения в организации яиц хордовых Для Spiralia характерно развитие, при котором отдельные бластомеры на ранних стадиях дробления уже запрограммированы к дифференцировке в определенном направлении, не изменяющемся даже в том случае, если изолировать их от
1. ЕСТЕСТВЕННАЯ АГРЕССИВНОСТЬ И ЭВОЛЮЦИОННЫЕ МЕХАНИЗМЫ, ЕЕ ОГРАНИЧИВАЮЩИЕ
1. ЕСТЕСТВЕННАЯ АГРЕССИВНОСТЬ И ЭВОЛЮЦИОННЫЕ МЕХАНИЗМЫ, ЕЕ ОГРАНИЧИВАЮЩИЕ Три закона робототехники1. Робот не может причинить вред человеку или своим бездействием допустить, чтобы человеку был причинен вред.2. Робот должен повиноваться всем приказам, которые отдает
1.3. Существование самоотверженности и ее эволюционные преимущества
1.3. Существование самоотверженности и ее эволюционные преимущества Логике примитивного социал-дарвинизма противоречат факты массового героизма и самоотвержения, с существованием героической верности долгу, с самоуничтожением ради выполнения долга, с существованием
Когда-то наши эволюционные пути разошлись
Когда-то наши эволюционные пути разошлись Человекообразные в области сексуальных и брачных отношений явно ушли от общих с человеком предков своими особыми путями. Общее с человеком у них только наличие менструальных циклов, заменивших овуляцию раз в год, характерную
Звенья одной цепи
Звенья одной цепи В. П. Данилин решил узнать, как люди оценивают интервалы времени, прошедшие во сне. Он исходил из предположения, что адекватная оценка времени означает во всех случаях, что у человека в памяти фиксируется непрерывная последовательность событий,
Эволюционные предпосылки гомосексуальности
Эволюционные предпосылки гомосексуальности Гомосексуальное поведение — явление отнюдь не сугубо человеческое. Оно широко распространено у животных в естественной среде обитания. Гомосексуальные отношения можно часто наблюдать в группах холостяков у павианов
Эволюционные корни ритуала
Эволюционные корни ритуала Антропологи уже давно пришли к выводу, что ритуалы в древнейших человеческих сообществах способствовали выживанию, поддерживая в данном клане или племени чувство особой идентичности и общей судьбы[91]. Ритуал постоянно напоминал членам