Соотношение между гомеозисом и нормальной сегментацией
Соотношение между гомеозисом и нормальной сегментацией
В детерминации индивидуальности сегментов, по-видимому, непосредственно участвуют две группы гомеозисных локусов. Интересно указать, что эти два набора генов представляют собой два кластера тесно сцепленных между собой локусов, находящихся в правом плече третьей хромосомы. О членах группы, расположенной дистальнее, мы уже говорили. Она известна под общим названием Bithorax Complex, и в нее входит не менее 10 комплементарных локусов. В другую, более проксимальную группу входит не менее 6 членов, и она известна под названием Antennapedia Complex. Связи этих двух комплексов друг с другом и с другими гомеозисными локусами приведены на рис. 8-5. На рис. 8-8 показан состав каждого комплекса и его местоположение в политенных хромосомах. Члены группы ВХ-С ответственны за детерминирование индивидуальности каждого из сегментов, расположенных кзади от сред-негрудного сегмента у личинки и у имаго. Как мы уже видели, в отсутствие локусов bx и pbx заднегрудь имаго превращается в среднегрудь. Мутация bxd (bithoraxoid) вызывает сходное превращение первого брюшного сегмента в среднегрудной. В этом случае трансформация наблюдается также и у личинки. Эти три рецессивные мутации комплементарны друг другу (т. е. особи с генотипом bx/pbx или bx/bxd, например, обладают нормальным фенотипом, но ни одна из трех не комплементарна доминантной мутации Ubx (Ultrabithorax). Льюис (Lewis) пришел к выводу, что дефекты, вызываемые мутацией Ubx, соответствуют мутациям в той части комплекса ВХ-С, которая ответственна за регуляцию локусов bx, pbx и bxd. Особи, гомозиготные по какой-либо мутации Ubx, гибнут на поздних стадиях зародышевого развития; при этом у них происходит превращение третьего грудного и первого брюшного сегментов во вторую грудь - результат, соответствующий представлениям Льюиса о функции локуса Ubx. Чем больше дистальных локусов комплекса ВХ-С подвергается делеции (рис. 8-8, Б), тем больше брюшных сегментов превращается в грудные сегменты. Так делеция локусов iab-2, iab-3 и iab-8 (рис. 8-8, Б) приводит к превращению брюшных сегментов в среднегрудь. При делеции всего комплекса ВХ-С все сегменты от заднегрудного до 8-го брюшного превращаются в среднегрудь, как это схематически показано на рис. 8-9. Все эти превращения - результат отсутствия функции генов. Однако у комплекса ВХ-С есть и другие доминантные варианты, приводящие к аномальной активации функций в пределах этого комплекса. Доминантная мутация Cbx (Contrabithorax) приводит к развитию мух, крылья которых превращены в жужжальца. Как показала генетическая проверка, это вызвано дерепрессией функций локусов bx+ и pbx+ в среднегруди, где они обычно не проявляют активности. Такого рода мутации существуют по крайней мере в трех точках комплекса ВХ-С и, согласно предположениям, оказывают воздействие на cis-регуляторный участок, контролирующий действие входящих в этот комплекс локусов, которое в норме ограничено отдельными сегментами.
Комплекс гомеозисных локусов, расположенный более проксимально (ANT-C, рис. 8-8, A), вызывает сходные превращения сегментов в передних частях зародыша и имаго. Локус pb (proboscipedia) необходим для нормального развития нижнегубного сегмента взрослой особи. В отсутствие функции pb+ нижнегубные щупики имаго превращаются в переднегрудные ноги. Делеция локуса Scr (Sex combs reduced) приводит к превращению переднегруди в среднегрудь. Это лучше всего видно на взрослых самцах дрозофилы, у которых половой гребешок (структура, обычно имеющаяся только на переднегрудных ногах) отсутствует. Кроме того, о превращении первой пары ног во вторую свидетельствует общая картина распределения щетинок на переднегрудных ногах как у самцов, так и у самок. Такое же превращение можно наблюдать у личинок, у которых типичная для переднегруди морфология изменяется, так что переднегрудь становится похожей на среднегрудь. Наконец, некоторые аллели локуса Antp обусловливают превращение среднегруди и заднегруди в переднегрудь как у личинок, так и у имаго. Это можно наблюдать у взрослых самцов, у которых при этом появляются половые гребешки на всех шести ногах. При делеции всего комплекса ANT-C зародыш гибнет на поздней стадии развития, перед самым вылуплением личинки первого возраста. Изучение этих особей, а также более ранних стадий до наступления гибели показало, что все их сегменты от максиллярного до заднегрудного похожи на переднегрудные сегменты. Проявление этих двух последних нарушений у зародыша представлено на рис. 8-9.
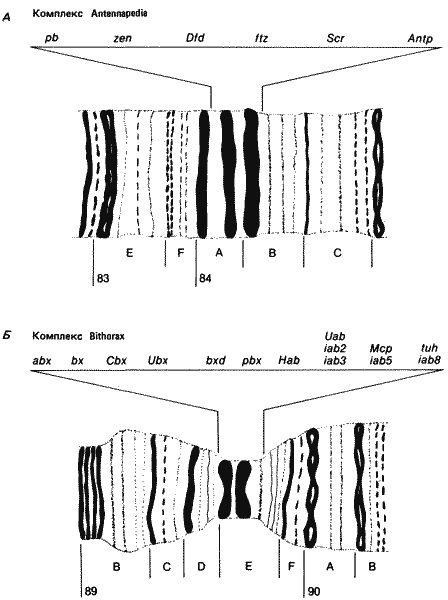
Рис. 8-8. Схематическое изображение участков политенных хромосом Drosophila melanogaster, в которых локализованы комплексы ANT-C(A) и ВХ-С(Б). Над каждым участком указаны локусы, входящие в эти комплексы, pb - proboscipedia; zen - zerknult; Dfd - Deformed; ftz - fushitarazu; Scr - Sex combs reduced; Antp - Antennapedia; abx - anteriobithorax; bx - bithorax; Cbx - Contrabithorax; Ubx - Ultrabithorax; bxd - bithoraxoid; pbx - postbithorax; Hab - Hyperabdominal; Uab - Ultraabdominal; Мер - Miscadestral pigmentation; tuh - tumorous head; iab - infraabdominal.
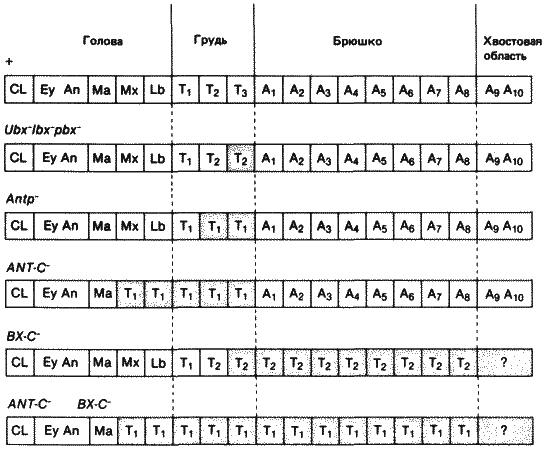
Рис. 8-9. Схема превращений сегментов, наблюдаемых после частичных или полных делеций комплексов ANT-C и ВХ-С. Трансформированные сегменты отмечены пунктиром, и на них указано, в какие сегменты они превратились. Слева, над каждой схемой-различные генотипы. Показанные здесь трансформации представляют собой упрощенную компиляцию эффектов, оказываемых этими мутациями на взрослых особей, личинок и зародышей. Однако в большинстве случаев лишь предполагается, что эти трансформации имеются у зародыша в начальный момент, когда отдельные сегменты приобретают свою индивидуальность. Еу - глаз; An - антенна; остальные обозначения, те же что на рис. 8-6.
Подобно комплексу ВХ-С, доминантное усиление функции происходит также в результате мутаций, относящихся к комплексу ANT-C. Наиболее очевидное из них - нарушение функции локуса Antp, вызывающее превращение антенны в ногу. Функция аллеля Antp+, установленная на основе фенотипа, получающегося при делеции этого локуса, заключается в обеспечении надлежащего развития средне- и заднегруди. При доминантных мутациях в локусе Antp эта функция проявляется в передних члениках антенны и в ее зачатках. Аллель Antp+ не участвует в нормальном развитии антенны, о чем свидетельствует нормальная морфология антенны, даже в том случае, если в этом органе с помощью соматического кроссинговера создаются гомозиготные клоны клеток Antp-. Если же создать такие клоны во второй и третьей паре ног, то развитие пойдет по типу переднегруди или антенны. Таким образом, функция аллеля Antp+, очевидно, необходима для нормального развития вентральных частей средне- и заднегруди, а превращение антенны в ногу происходит в результате аномальной активации этой функции в зачатке антенны.
При делеции как комплекса ВХ-С, так и комплекса ANT-C получается картина сегментации, изображенная в нижнем ряду рис. 8-9: голова зародышей образована тремя сегментами, за которыми следует ряд одинаковых сегментов, похожих на переднегрудные. Такие зародыши гибнут. Следовательно, гены, входящие в эти два комплекса, необходимы для определения особой судьбы большинства различных гнатоцефалических, грудных и брюшных сегментов. Отдельные локусы функционируют в определенных сегментах, причем эта функция не выходит за пределы отдельных сегментов. Трансформации сегментов, вызываемые делецией генов, входящих в комплексы ВХ-С или ANT-C, или мутациями этих генов, не сводятся лишь к поверхностному изменению внешних кутикулярных структур. Если извлечь из личинок имагинальные диски, диссоциировать их и допустить реагрегацию клеток, то клетки одинаковых дисков соединяются друг с другом, а клетки из разных дисков отделяются одни от других. Если, однако, антенны трансформированы в результате гомеозиса в ногу, то эти клетки антенны-ноги воссоединяются и совместно образуют нормальные бластемы ноги. Эти результаты указывают на то, что поверхностные свойства клеток и их способности к распознаванию друг друга изменились под действием мутации.
С помощью метода соматической рекомбинации была сделана попытка оценить число клеток, из которых на стадии клеточной бластодермы состоят различные имагинальные диски в момент их детерминации. Для диска, из которого образуются жужжальца, их число равно примерно 10, а для диска крыла - около 20. Кроме того, удалось проследить за динамикой пролиферации этих двух дисков, получая рекомбинантные клоны их клеток на более поздних стадиях развития. С помощью этого метода Гарсиа-Беллидо (Garcia-Bellido) и его сотрудникам удалось показать, что у двойных мутантов bx pbx изменяется как динамика роста, так и число клеток, вовлеченных в диск жужжальца на стадии клеточной бластодермы. Оба этих свойства, подобно морфологии диска, изменяются, и жужжальца становятся похожими на крыло, т.е. на производное среднегруди. То обстоятельство, что программа развития как личинки, так и имагинальных дисков при гомеозисе изменяется на очень ранней стадии (для зачатков, по-видимому, на стадии клеточной бластодермы), можно считать указанием на важную роль гомеозисных генов в фактической детерминации судьбы клеток. В пользу такого утверждения свидетельствует возможность получить фенокопии трансформаций, типичных для мутантов bithorax, путем нагревания зародышей, находящихся на стадии клеточной бластодермы. У зародышей в возрасте 2 ч, подвергнутых температурному шоку, возникают ненаследственные изменения развития, сходные с изменениями, наблюдаемыми у мутантов bx. Как показал Гарсиа-Беллидо, к таким же результатам приводит обработка зародышей того же возраста эфиром. Эффективность такой обработки значительно возрастает, если изменить дозу генов комплекса ВХ-С посредством гетерозиготных делеций bx и pbx; при этом частота фенокопии повышается. Эти наблюдения являются убедительным доводом в пользу того, что мишенью, на которую направлено действие гомеозисных локусов, служат ранние процессы детерминации. Эти локусы необходимы, однако, и для поддержания такого состояния. Температурочувствительные мутации в локусах bx и pbx обладают температурочувствительным периодом, соответствующим третьей личиночной стадии (спустя 4-5 дней после детерминации клеток имагинальных дисков). Таким образом, характер детерминации этих клеток можно изменить после его первоначального формирования, но до дифференцировки, происходящей в процессе метаморфоза.
Результаты генетических и эмбриологических исследований гомеозисных генных комплексов Antennapedia и bithorax показывают, что эти локусы действительно играют роль переключателей, определяющих судьбу отдельных сегментов. Переключение происходит в период детерминации и, по-видимому, представляет собой результат позитивного действия названных локусов этих комплексов на батареи других генов. Следует отметить, что альтернативный путь развития, происходящий в отсутствие этих генов, не бывает хаотическим; в норме он наблюдается у некоторых других частей животного, так что эти гены можно в самом деле рассматривать как переключатели, производящие выбор из нескольких альтернативных состояний. Изменение реакции, вызванное гомеозисной мутацией, не приводит, как хотелось бы Гольдшмидту, к появлению «перспективных монстров», обладающих потенциальными возможностями для макроэволюционных событий. На самом деле изменения эти носят атавистический характер и могут дать сведения об истории генетических регуляторных механизмов, приобретенных в процессе эволюции. Вероятно, особенно ясно это можно продемонстрировать на примере эволюционной истории насекомых.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
4. Взаимоотношения между клетками
4. Взаимоотношения между клетками О взаимоотношениях между клетками в морфогенезе мы говорили и в предыдущих разделах этой главы. Изменения формы клеток, их движение и другие виды поведения в организме всегда осуществляются совместно многими клетками и в зависимости их
Перекресты. Расстояния между генами
Перекресты. Расстояния между генами Обычно дрозофилы сероватые, но есть среди них «блондины»—золотисто-желтые. Это рецессивный признак, вызванный геном из первой хромосомы. Его обозначают буквой «у», от английского слова yellow — желтый. Раз есть рецессив, значит, есть и
МЕЖДУ ДВУХ СТУЛЬЕВ
МЕЖДУ ДВУХ СТУЛЬЕВ Но вот как раз заслуг-то эволюция никогда и не помнит. Основным же просчетом бывшего царя природы оказался слишком незначительный размах его владений. Узкие прибрежные полосы — вот и всё. Верность воде в конце концов и привела стегоцефалов к упадку. А
Соотношение объемов талии и бедер
Соотношение объемов талии и бедер Правило сбалансированности, или «золотой середины», относится и к другим пропорциям тела.Психолог Дивендра Син продемонстрировала группе американских мужчин серию рисунков, изображающих молодых женщин, и поинтересовалась, какой тип
Конкуренция между терапевтами и узкими специалистами — борьба между коллегами
Конкуренция между терапевтами и узкими специалистами — борьба между коллегами Отношения между терапевтами и специалистами стали все более и более напряженными. Процент вторых по отношению к первым имеет тенденцию к росту в направлении частной практики. Специалисты в
Сговор между государством и лабораториями
Сговор между государством и лабораториями В начале XIX в. богиня Разума отказалась от ранее накопленного медицинского опыта. Во имя революции, в основном материалистической и механической, природа, которая не была продуктом человеческого разума, была отодвинута на
2.1. Вопрос нормы и патологии. Определение нормальной психики
2.1. Вопрос нормы и патологии. Определение нормальной психики В работе с животными, проявляющими аномальное поведение, одним из базовых понятий, на которое должны опираться в своей деятельности специалисты в области поведенческой медицины, это различение нормы и
II. Между песком и небом
II. Между песком и небом Способы использования пространства и других условий окружающей среды жителями континентального шельфа отличаются удивительным разнообразием. Многие из них, выйдя из личиночного состояния и осев на одном месте, передвигаются на расстояния,
Соотношение потребностей
Соотношение потребностей Первое, что вспоминается, – это «пирамида Маслоу». В ее основании лежат потребности в еде и питье, а на самой вершине находится идеальная потребность, которую Абрахам Маслоу назвал потребностью в самореализации.В первом приближении такая схема
Соотношение половых хромосом и аутосом как фактор определения пола
Соотношение половых хромосом и аутосом как фактор определения пола Ученые уже в первой половине XX века активно обсуждали вопрос о том, что же лежит в основе определения пола. Широкую известность получила балансовая теория К. Бриджеса. По мнению американского генетика, не
Соотношение полов в популяции
Соотношение полов в популяции Как показывают исследования зоологов, для большинства видов животных типичным является соотношение самцов и самок в популяции близкое 1:1. В норме у человека на 100 новорожденных девочек приходится 106 мальчиков. Так что и у человека
Соотношение талии к бедрам
Соотношение талии к бедрам Теперь следует вспомнить, что кроме лиц есть еще и фигуры. Хорошо известно, какой огромной притягательной силой обладают пропорции тела женщины. Над пресловутым «90-60-90» многие посмеиваются, однако этологи показали, что это соотношение возникло
Соотношение бюста к талии
Соотношение бюста к талии До сих пор мы говорили об отношении талии к бедрам. А как обстоит дело со второй частью «магической» пропорции — отношением бюста к талии? Насколько оно стабильно в эпохальном и кросс-культурном аспекте? Б. Сильверстен с коллегами
Соотношение плеч к бедрам и мужская привлекательность
Соотношение плеч к бедрам и мужская привлекательность Как следует из предыдущих разделов, часть морфологических признаков человека демонстрирует выраженный половой диморфизм и сопряжена с репродуктивными качествами носителя. Такие признаки являлись и являются