Становление пространственной организации и полярные координаты
Становление пространственной организации и полярные координаты
Хадорн (Hadorn) и его ученики обнаружили, что если у личинки третьего возраста дрозофилы удалить один из имагинальных дисков и пересадить его путем инъекции новой личинке-реципиенту, то он будет дифференцироваться вместе с этим реципиентом. Более того, если диск разрезать на части, то каждый его фрагмент самостоятельно дифференцируется, образуя часть тех структур, которые образовал бы этот диск при нормальных обстоятельствах, причем из каждого отдельного фрагмента развиваются специфичные для него структуры. Часть такого рода карты для диска крыла представлена на рис. 9-4. Эти эксперименты показали также, что в имагинальных дисках не происходит регуляции типа морфоллаксиса. У них наблюдается, однако, эпиморфоз. Если фрагменты имагинального диска поместить не в личинку, а в брюшко взрослой самки дрозофилы, то в результате клеточных делений они растут, но не дифференцируются. Если выращивать фрагменты на протяжении недели, а затем пересадить их в метаморфизирующую личинку, то они дифференцируются, образуя вполне согласованные структуры. Если разрезать диск на две неравные части, то из большего фрагмента дифференцируются и те структуры, которые в норме образуются из клеток меньшего фрагмента. Что касается меньшего фрагмента, то он образует собственный зеркально-симметричный дубликат. Результаты двух таких экспериментов схематически изображены на рис. 9-4.
Можно представить себе, что положение клеток в пространстве определяется набором радиальных и угловых координат, причем положение каждой клетки задается особой парой сигналов. Если имагинальный диск разрезать на две части, то в результате смыкания поверхностей разреза в каждой части приходят в соприкосновение такие позиционные значения, которые обычно не примыкают друг к другу. При наличии подходящих условий (например, при выращивании в брюшке взрослой самки) это соприкосновение индуцирует рост и клеточное деление. Рост, согласно модели, возникнет в том случае, если различные угловые либо радиальные значения позиционной информации окажутся в непосредственном контакте. Клетки, образующиеся в результате этого процесса, воссоздадут недостающую позиционную информацию в общем таким же образом, как прогрессивная зона моделирует почку конечности цыпленка. Чтобы объяснить результаты, наблюдаемые при разрезании имагинального диска на два неравных фрагмента, Френч, Брайант и Брайант сформулировали правило кратчайшей интеркаляции. Его легче всего понять на примере, приведенном на рис. 9-4, В. Маленький фрагмент интеркалирует (восстанавливает) позиционные значения 4 и 5, потому что это кратчайший путь по отношению к двум сохранившимся на данном фрагменте угловым значениям (6 и 3). Концептуально эта модель сходна с моделью линейного градиента, в которой данный участок градиента способен регенерировать только более низкие позиционные значения. Большой фрагмент интеркалирует ту же самую недостающую информацию (позиционные значения 4 и 5 на окружности), но тем самым он восстанавливает полную окружность и создает всю необходимую позиционную информацию. Маленький фрагмент дуплицируется, а большой регенерирует; это и дает требуемый экспериментальный результат. Необходимо ввести еще одно правило - правило полной окружности; согласно этому правилу, для создания дистальных структур (участки, специфицируемые в центре круга) необходимо регенерировать все угловые позиционные значения. Из этого последнего правила есть несколько исключений. Однако они не умаляют фундаментальной полезности данной модели как способа представить себе, каким образом процесс онтогенеза извлекает пространственную организацию из видимого отсутствия формы. При этом не следует забывать, что такие модели представляют собой абстракции очень сложных клеточных и генетических событий и в лучшем случае могут дать лишь приближенное представление о процессах, происходящих в действительности. Необходимо постоянно помнить слова Уоддингтона о том, что «...эмбриональное поле - это, в сущности, концепция, пригодная в сфере рассуждений, относящихся к многомерным пространствам. Любую попытку свести ее к трем или даже четырем измерениям плюс одна переменная поля следует рассматривать как решительное абстрактное упрощение, которое может быть оправдано для некоторых особых целей, но к которому всегда следует относиться с большой осторожностью».
Брайант и его сотрудники обнаружили, что позиционная информация, содержащаяся в имагинальных дисках, по-видимому, одинакова во всех дисках. У тех фрагментов дисков крыла, которые при нормальных условиях дуплицируются, можно индуцировать регенерацию, культивируя их совместно с реципрокными фрагментами, убитыми облучением. Убитые фрагменты не поставляют в регенерирующую структуру живые клетки, но, по-видимому, могут поставлять ей позиционную информацию. Информация эта может поступать, очевидно, не только от данного, но и от любого другого диска.
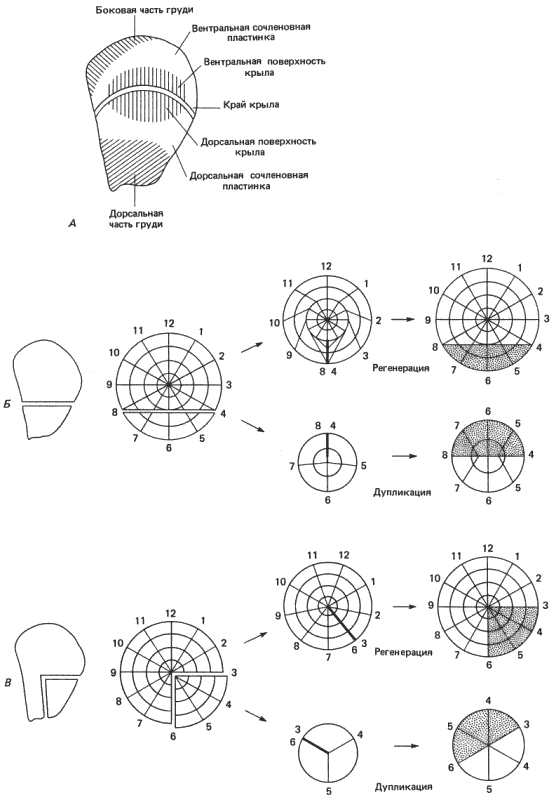
Рис. 9-4. Объяснение результатов регенерации и дупликации диска крыла у Drosophila melanogaster с помощью модели полярных координат. А. Примерная карта зачатков диска крыла. Б и В. Слева - два вида разреза этого диска. В обоих случаях меньший фрагмент дуплицируется, а в большем происходит регенерация недостающих элементов. Результат представлен справа от каждого разрезанного диска в терминах модели полярных координат. Новообразованные секторы показаны точками. Правила регенерации, соответствующие этой модели, изложены в тексте. (French, Bryant, Bryant, 1976.)
Приведем конкретный пример: проксимальный дуплицирующийся фрагмент диска крыла, подобный изображенному на рис. 9-4, А, можно индуцировать к регенерации дистальных структур крыла, выращивая его в присутствии убитых дистальных фрагментов диска крыла или диска жужжальца. Этот результат согласуется с данными о том, что гомеозисные мутации вызывают трансформации, которые сериально гомологичны, т.е. дистальные части антенны превращаются в дистальные части ноги. Отсюда следует, что позиционная информация во всех имагинальных дисках одинакова; различие состоит в том, как она интерпретируется. Было показано, что эта очевидная универсальность позиционной информации выходит даже за видовые границы. Тикле (Tickle) и его сотрудники показали, что ЗПА, взятая из задней части почки конечности мыши, способна передать информацию, необходимую для установления положения передне-задней оси конечности у цыпленка. Пересадив ЗПА мыши на передний край почки конечности цыпленка, можно индуцировать развитие у цыпленка добавочных пальцев. Сходный результат получили Фаллон и Кросби (Fallon, Crosby), используя ЗПА из зародышей черепахи. Поэтому создается впечатление, что информация, специфицирующая поле развивающейся конечности у позвоночных, в процессе эволюции оставалась неизменной, а изменения коснулись реакции клеток на эту информацию. Характер этой реакции почти наверное определяется различиями в генной экспрессии или, если говорить прямо, во включении генетических переключателей.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
2.1. Становление и основные положения синтетической теории эволюции
2.1. Становление и основные положения синтетической теории эволюции Эволюционизм возник как альтернатива учению о неизменности видов. Вопросы, связанные с возникновением и развитием жизни, прошли через всю интеллектуальную историю человечества. Количество литературы,
3.2. Становление этологии и вопросы эволюции поведения
3.2. Становление этологии и вопросы эволюции поведения Этология возникла в середине 1930-х гг. как наука, изучающая поведение животных в естественной среде обитания. Эта наука дала миру целую плеяду талантливых ученых, однако даже на таком ярком фоне выделяются имена ее
4.1. Типы социальной организации
4.1. Типы социальной организации В этологических исследованиях был обнаружен широкий диапазон различных видов социальной организации. У высших животных их социальные структуры и социальное поведение в процессе эволюции достигают исключительной степени сложности и
10.1. Становление взглядов на проблему когнитивных способностей животных
10.1. Становление взглядов на проблему когнитивных способностей животных Первые систематизированные данные по исследованию когнитивных способностей животных получил английский ученый, ученик Ч. Дарвина Дж. Романес (1848–1894). В 1883 г. выходит его книга «Интеллект животных».
11.1. Становление психологии
11.1. Становление психологии В дословном переводе «психология» означает «наука о душе». В научной терминологии вместо слова «душа» стали употреблять слово «психика», что, однако, не прибавило определенности этому понятию. Сложность, неопределенность, «неуловимость»
Становление локализации и пространственной организации
Становление локализации и пространственной организации Во время оогенеза происходит чрезвычайно активная транскрипция генов и накопление ооцитами мРНК. Накапливающиеся в ооцитах мРНК столь разнообразны по своим последовательностям, что, как только после начала
Эволюционные изменения в организации яиц хордовых
Эволюционные изменения в организации яиц хордовых Для Spiralia характерно развитие, при котором отдельные бластомеры на ранних стадиях дробления уже запрограммированы к дифференцировке в определенном направлении, не изменяющемся даже в том случае, если изолировать их от
Глава 9 Становление пространственной организации
Глава 9 Становление пространственной организации Пространственная организация и происхождение формы Главная проблема, стоящая перед биологами, занимающимися изучением развития, - объяснить механику процессов, в результате которых из одноклеточной зиготы образуется
Становление пространственной организации и спецификация местоположения
Становление пространственной организации и спецификация местоположения Презумптивную область ноги, которую пересаживал Гаррисон, можно рассматривать как эмбриональное поле. В более общем смысле поле - это участок зародыша или группа клеток, в пределах которых может
4.2. Возникновение клеточной организации
4.2. Возникновение клеточной организации Учеными было доказано, что после испарения воды из реакционного объема в амфифильных липидоподобных и липидных молекулах формируются жидкокристаллические агрегаты, в которых молекулы расположены периодическими слоями, как в
Становление ученого
Становление ученого Его жизнь прошла в беспрерывной борьбе и беспримерных дерзаниях. Десятилетиями длилась эта борьба. Трудно было поверить, что он обнаружит так много сил, найдет в себе столько решимости. Безудержная мысль Александра Васильевича Вишневского и на