Гибель клеток в процессе нормального развития
Гибель клеток в процессе нормального развития
Дегенерация вольфовых протоков в процессе развития особей женского пола и мюллеровых каналов у особей мужского пола - примеры наличия структур, которые вначале развиваются, а затем подвергаются некрозу. Эти процессы, подобно всем другим рассматриваемым здесь онтогенетическим событиям, находятся под контролем генов. Специфическим примером служит развитие конечностей у четвероногих. У цыпленка почки конечностей закладываются в виде латеральных утолщений соматоплевры примерно на 55-м часу развития. Эти почки вырастают из тела зародыша в виде выступов, покрытых эктодермой и заполненных мезодермальной тканью. По мере продолжения роста начинают проступать контуры конечности, будь то крыло или нога. Процесс формирования контуров сопровождается отмиранием клеток в ряде мезодермальных участков конечности. На ранних стадиях образования почки конечности участки некроза можно выявить с помощью некоторых прижизненных красителей; такие участки есть и на переднем, и на заднем краях почки, там, где она соединяется со стенкой тела, а также в центре почки. Эти три участка называются соответственно передней и задней некротическими зонами и темными пятнами (рис. 7-16). Передняя и задняя зоны обеспечивают формирование контуров проксимальных областей конечности, а темное пятно - отделение большой берцовой кости от малой в ноге и лучевой от локтевой - в крыле. Происходящее на более поздних стадиях развития конечности разделение пальцев сопровождается некрозом межпальцевых участков (рис. 7-16).
Сондерс и Фаллон (Saunders, Fallen), изучая заднюю некротическую зону (ЗНЗ) почки крыла цыпленка, выявили несколько интересных особенностей этой группы клеток. Максимальной протяженности ЗНЗ достигает на 96-м часу развития; к этому времени гибнет 1500-2000 клеток. Эти клетки фагоцитируются популяцией из почти 150 макрофагов. Если мезодерму проспективной ЗНЗ извлечь из почки крыла за 40 ч до того, как становится заметным некроз, и пересадить ее на боковую поверхность тела реципиента, она все равно гибнет и притом в те же сроки. Сроки некротизации и гибели заложены в самих клетках ЗНЗ и не зависят от возраста реципиента. Однако мезодермальные клетки, находящиеся в почке конечности, которая примыкает к эксплантированной ЗНЗ, не погибают. Поэтому можно думать, что клетки ЗНЗ обладают механизмом, получившим название «внутренние часы смерти». Эти часы запускают механизм гибели клеток на очень ранних стадиях развития конечности, после чего клетки идут по предначертанному им пути. Ход этих часов, однако, до некоторой степени обратим. Если ЗНЗ пересадить на дорсальную поверхность почки крыла, то ее клетки не гибнут. Причину этого изучали, выращивая клетки ЗНЗ вместе с различными другими фрагментами мезодермы. Выращивание мезодермы из ЗНЗ с соматической мезодермой приводит к такому же результату, как пересадка ее in vitro на боковую поверхность зародыша-реципиента: клетки ЗНЗ гибнут в надлежащие сроки. Если, однако, ЗНЗ культивировать с дорсальной мезодермой крыла, то она не гибнет. Создается впечатление, что мезодерма из ненекротической области крыла защищает ЗНЗ от гибели, останавливая «часы смерти». Это было подтверждено экспериментами, в которых клетки ЗНЗ сначала пересаживали на дорсальную поверхность почки крыла, а затем лишали их защитных воздействий дорсальной мезодермы крыла, удаляя их с места пересадки через разные сроки после того, как по «часам смерти» они должны были бы погибнуть. Будучи удалены с места пересадки, эти клетки погибали; выживали только те из них, которых содержали in vitro в условиях защиты не менее 6 дней. К этому времени «часы» совершенно останавливались и клетки ЗНЗ выживали, даже в отсутствие дорсальной мезодермы крыла. Таким образом, гибель клеток, происходящая в проксимальных частях крыла, представляет собой результат действия двух факторов: неких независимых от остального организма клеточных часов, запускаемых в определенной группе клеток, и местоположения этих клеток относительно остальной мезодермы конечности.
Анализ гибели клеток в межпальцевых участках позволил выявить еще один уровень регуляции. Как видно из рис. 7-16, гибель клеток в этих участках лапы утки невелика, и в результате между вторым, третьим и четвертым пальцами образуются перепонки. В промежутке между первым и вторым пальцами, где происходит некроз, перепонки нет. Сондерс и Фаллон создавали химеры, соединяя мезодерму из почки ноги утки с эктодермой цыпленка и наоборот. Такие гибридные почки ноги они пересаживали затем на боковую поверхность тела куриного зародыша, служившего реципиентом, и наблюдали за некрозом межпальцевых клеток. В обоих случаях картина некроза соответствовала тому, что характерно для утки. Отсюда можно сделать вывод, что утиная эктодерма, подобно мезодерме дорсальной поверхности крыла, может подавлять некроз, наблюдаемый обычно при развитии ноги цыпленка. Однако эктодерма куриного зародыша не индуцирует некроз в утиной мезодерме. Возможно, поэтому, что регуляция некроза в межпальцевых участках происходит таким же образом, как в ЗНЗ; т.е. что это автономная особенность межпальцевой мезодермы, которая может проявиться или не проявиться в зависимости от среды, в которой окажутся презумптивные некротические клетки.
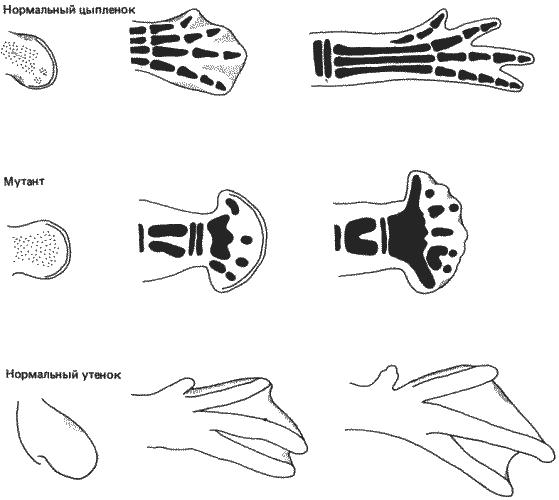
Рис. 7-16. Развитие задней конечности цыпленка и утки. Участки гибели клеток на задних и передних краях ноги зародышей, а также некроз в межпальцевых участках у зародышей цыпленка (но не утки) показаны точками. Мутация talpid, по-видимому, элиминирует некроз на большей части участков, что приводит к образованию широкой веслообразной конечности со сросшимися пальцами, схематически изображенной в центре рисунка. Темные участки на нормальной и мутантной конечности цыпленка соответствуют участкам образования хряща (Saunders Jr., Fallen, 1966; Hinchliffe, Thorogood 1974; с изменениями).
Хинчлифф и Торогуд (Hinchliffe, Thorogood), проведя анализ мутации ta (talpid) у цыпленка, показали, что гибель клеток в процессе развития конечности действительно находится под генетическим контролем. Мутация ta наследуется как простой аутосомный рецессивный признак и в гомозиготном состоянии обусловливает костную полидактилию и синдактилию мягких тканей в крыле и ноге. Полидактилия возникает в результате слияния лучевой и локтевой костей и большой и малой берцовых костей. Образующиеся при этом конечности, особенно нога, широкие и веслообразные, и на них бывает по 6-7 пальцев вместо четырех нормальных (рис. 7-16). Для развития этих конечностей характерно почти полное отсутствие гибели клеток. Ширина мутантной лапы объясняется отсутствием передней и задней некротических зон, поскольку при этом не происходит определения контуров, которые у нормальных особей сужают эту часть конечности. Слияние костей предплечья или голени - результат отсутствия темного пятна. Это несостоявшееся разделение костей может быть также результатом расширения дистальных частей конечности и кажущейся зеркальной симметрии ее строения (рис. 7-16). Кроме того, у мутантов отсутствуют межпальцевые некротические зоны, имеющиеся в нормально развивающейся конечности, и в дальнейшем пальцы у них не отделяются друг от друга. Интересно отметить, что синдактилию сходного типа наблюдал Джонсон (Johnson) у мутантных мышей polysyndactylous, но без костной полидактилии. Остается еще установить, оказывает ли мутация talpid воздействие на «часы смерти» или же она дает возможность «спасающему фактору», продуцируемому дорсальной мезодермой, достигнуть обреченных на гибель клеток. Какой бы ни была непосредственная причина, ясно, что процесс некроза клеток регулируется генетически и что в создании дефинитивной морфологии конечности позвоночных гибель клеток играет важную роль. В связи с этим последним моментом следует указать, как это делают Хинчлифф и Торогуд, что простые раздвоенные структуры, наблюдаемые в конечностях мутантов talpid, напоминают по своему характеру элементы, имеющиеся в плавниках древних кистеперых рыб, таких как Eusthenopteron и Sauripterus. Возможно, поэтому, что сложный тип развития конечностей, обнаруженный у высших четвероногих позвоночных, мог возникнуть в результате процесса клеточного некроза, регулируемого и моделируемого генетически.
Существуют также мутации, расширяющие зоны некроза. Такие мутации дрозофилы, как Bar (полосчатые глаза) или vestigial (зачаточные крылья), сильно уменьшающие глаза или крылья, действуют путем увеличения участков гибели клеток, наличие которых составляет один из элементов нормального развития этих двух структур. Мутации wingless и rumpless у кур действуют, по-видимому, при помощи сходного механизма, создающего возможность для увеличения зоны некроза. Можно представить себе, что изменения этого типа (в менее резко выраженной форме) играют важную роль в редукции или элиминации структур, необходимых на одной стадии развития, но ненужных на более поздней стадии. Простым примером служит хвост у головастиков лягушек. В этом случае, как и в случае мутаций, подавляющих гибель клеток, существуют два возможных уровня регуляции процесса элиминации хвоста: внутренние часы смерти могут активизироваться в большем числе клеток или же внешние факторы, такие например как способное к диффузии защитное вещество, содержащееся в дорсальной мезодерме крыла, могут быть элиминированы.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Обреченные на гибель
Обреченные на гибель Теперь вы понимаете, почему иммунитет, спасающий нас от смерти в борьбе с микробами, является в других случаях нашим врагом.Врагом, конечно, относительным. Скажем мягче: иммунитет в некоторых случаях мешает. Иммунитет следит за постоянством
Роль нормального питания
Роль нормального питания Сообщения о том, что в передовых капиталистических странах широко распространено недоедание, поначалу были восприняты весьма скептически. Однако они подтвердились не только дальнейшими исследованиями, но и анализом воздействия на человека
Глава первая: Триумф и гибель динозавров
Глава первая: Триумф и гибель динозавров Следы исполинов О том, что на Земле когда-то жили исполины, перед которыми казался крошкою любой высокий человек, говорится в легендах и мифах самых разных народов. В ирландских сагах и сказках братьев Гримм, в Библии и преданиях
Роль полового опыта для нормального развития собаки
Роль полового опыта для нормального развития собаки Следующий практический вопрос, вызывающий споры в среде собаководов: насколько необходимо собаке для полноценного развития и здоровья приобретение полового опыта? Ответ зависит от пола животного.СукиДля суки важно
Дифференциальная экспрессия генов в процессе развития
Дифференциальная экспрессия генов в процессе развития Один из главных и общепризнанных догматов современной эмбриологии состоит в том, что, за исключением нескольких особых случаев, все клетки данного организма, независимо от того какими они становятся в
Гены, вступающие в действие на более поздних стадиях развития и в процессе роста
Гены, вступающие в действие на более поздних стадиях развития и в процессе роста Ясно, что мутации генов, непосредственно определяющих морфогенетические пути, в особенности тех генов, которые действуют на ранних стадиях, могут вызывать чрезвычайно резкие изменения
Глава 10 Адаптации экспрессии генов в процессе развития
Глава 10 Адаптации экспрессии генов в процессе развития Жизнь -это сила, которая проделывает бесчисленное множество экспериментов, пытаясь организовать себя ... мамонт и человек, мышь и мегатерий, мухи и отцы церкви - все это результаты более или менее успешных попыток
Рождение и гибель муравейника
Рождение и гибель муравейника Носильщики и «чемоданы». К муравейнику возле большого серого камня через дорогу тянется процессия муравьев. Муравьи переносят друг друга. Одни ползут с ношею домой, другие, порожние, спешат им навстречу. Поза переносимого муравья
Необыкновенная гибель шхуны «Перл»
Необыкновенная гибель шхуны «Перл» 10 мая 1874 года шхуна «Перл» вышла из Галле в Рангун и не пришла в порт. Она пропала без вести. В истории мореплавания зарегистрировано немало подобных случаев, можно назвать не один пропавший без вести корабль. Но гибель маленькой
Гибель зверьков
Гибель зверьков Забота о детях — великая идея, воплощенная эволюцией в реальность. Животные, которые ее еще не постигли и детей своих бросают на произвол судьбы, должны, чтобы не угас их род, плодиться часто и много, неэкономно выбрасывая, что называется, на ветер, как,
Гибель муравейника
Гибель муравейника Кто подолгу просиживал в лесу у муравейников, наблюдая за жизнью этих многочисленных и своеобразных насекомых, мог легко заметить, что муравейник от муравейника отличался не только внешним видом, но и поведением своих жителей. В молодых, небольших
5. Теория стадийного развития и особенности развития животных
5. Теория стадийного развития и особенности развития животных В основе управления развитием организмов лежит теория стадийного развития, которую сформулировал академик Т. Д. Лысенко, исходя из работы И. В. Мичурина и многочисленных собственных исследований.Несмотря на