Типы видообразования
Типы видообразования
До сих пор наше внимание было сосредоточено главным образом на скоростях молекулярной и морфологической эволюции, и нам удалось выбрать количественные показатели, приемлемые для нескольких эволюционных процессов. Однако такие показатели, как скорость изменения размеров в дарвинах или скорости эволюции ДНК в числе замен нуклеотидов за год, могут создать иллюзию непрерывности - градуализма - даже в тех случаях, когда на самом деле имел место прерывистый ряд событий. Если эволюция обычно происходит прерывистым образом, то возникает необходимость определить природу процесса, вызывающего быстрое и, возможно, радикальное эволюционное изменение. Согласно наиболее прочно укоренившемуся мнению, опирающемуся на популяционную биологию и генетику, решающую роль в эволюции играет видообразование.
В этом контексте вид определяется как группа скрещивающихся между собой организмов, имеющих общий генофонд. А от такого определения неотделимы процесс и механизм, с помощью которых происходит видообразование. Если члены данного вида имеют общий генофонд, то события, приводящие к разделению одного вида на два, должны выделять из этого генофонда отдельные части и препятствовать обмену генетической информацией между двумя отдельными популяциями. Поэтому нам необходимо выяснить природу механизмов, разделяющих зарождающиеся виды, - природу генетических изменений, их количество, необходимое для видообразования, и необходимое на это минимальное время. Однако, прежде чем очертя голову устремиться к этой нетронутой целине, следует сделать два предостережения. Как совершенно справедливо отметил Буш, никто никогда не наблюдал процесс видообразования от начала и до конца, так что само исследование видообразования представляет собой «науку ad hoc». To, что мы наблюдаем в природе, это только отдельные моменты, ряд отдельных кадров непрерывного процесса, и, располагая лишь этими мимолетными впечатлениями, мы вынуждены воссоздавать весь остальной процесс и лежащий в его основе механизм. Это несколько напоминает метод Шерлока Холмса, который приводил в изумление доктора Ватсона: все прошлое будущего клиента выводилось из того, как он хромает и какие курит сигары. Как мы сможем убедиться, процесс видообразования протекает у разных организмов по-разному, и любые относящиеся к нему обобщения даются с трудом. Справедливости ради следует упомянуть о том, что в литературе описано несколько случаев лабораторного «видообразования». Эти случаи по большей части возникали в результате экспериментального или интуитивного возведения преград, препятствовавших скрещиванию между особями, принадлежащими к одному и тому же виду. Остается выяснить, однако, в какой мере эти лабораторные события соответствуют тому, что происходит в природе.
Другое предостережение относится к различию между адаптивными изменениями в пределах данной популяции и разного рода изолирующими механизмами, вызывающими расщепление, или кладогенез. Каждая природная популяция обладает известным запасом изменчивости, будь то хромосомный, морфологический или биохимический полиморфизм. Можно также показать, что эти кариотипические, морфологические или ферментные признаки изменяются во времени, со сменой времен года, или в пространстве, например с высотой местности. Классическим примером адаптивного изменения этого типа служит индустриальный меланизм у пяденицы Biston betularia. В этом особом случае в течение XIX в. в результате развития промышленности и загрязнения среды угольной пылью и копотью в популяциях этой бабочки в центральных графствах Англии черная морфа стала преобладать над серой. Пяденицы днем отдыхают на стволах деревьев, потемневших от копоти, а поэтому птицы лучше различают на них серых бабочек и выедают их сильнее, чем черных. Конечно, в популяции произошло изменение, однако она при этом не распалась на две отдельные репродуктивно изолированные группы. Черные и серые бабочки продолжают спариваться и производят жизнеспособное плодовитое потомство. Другой пример - полиморфизм по хромосомным инверсиям у Drosophila pseudoobscura, так изящно проанализированный Добржанским и его учениками. В третьей хромосоме этих мух содержится много различных генных последовательностей, перестроенных по сравнению с произвольно выбранной стандартной последовательностью. Во многих локальных популяциях содержится по нескольку таких инвертированных последовательностей. Частота каждой данной последовательности в популяции изменяется, однако, на протяжении всего сезона, когда эти насекомые растут. Частота инверсий изменяется также с изменением высоты местности над уровнем моря, так что на разных высотах в популяции преобладают различные инверсии, т.е. наблюдается клинальная изменчивость их частоты. И в этом случае все мухи, несущие различные хромосомные последовательности, интерфертильны, а поэтому они не относятся к разным видам. Создается впечатление, что изменения частоты генов или структуры хромосом, происходящие в пределах одной популяции и не сопровождающиеся видообразованием, возможно, играют известную роль в поддержании адаптированности популяции, но не играют существенной роли в эволюционном процессе.
Границы видов, особенно видов животных, устанавливаются репродуктивной изоляцией между ними. Это разделение поддерживается разнообразными изолирующими механизмами, которые можно разбить на две широкие категории - презиготические и постзиготические, в зависимости от того, подавляется ли передача генетической информации до или после оплодотворения. Презиготические преграды служат для предотвращения слияния гамет и могут сводиться всего лишь к экологическим различиям между двумя предполагаемыми брачными партнерами. Если две группы животных экологически изолированы либо реальным физическим расстоянием, либо тем, что они занимают достаточно различные ниши в одной и той же общей области, то скрещивание между ними маловероятно. Второй тип наблюдаемой презиготической изоляции-это временная изоляция. Если животные различаются по суточным ритмам активности или если растения различаются по срокам цветения, то они лишены возможности обмениваться генетической информацией. Третий тип презиготической изоляции специфичен для полового процесса как такового. У многих животных выработались весьма сложные брачные церемонии, которые должны быть выполнены во всех деталях, для того чтобы могло совершиться спаривание и произошло слияние гамет. В некоторых случаях при этих церемониях происходит не только обмен слуховыми и зрительными сигналами, но также выделение самцом, самкой или обоими партнерами специфических феромонов или половых аттрактантов. Четвертый презиготический механизм заключается в физической несовместимости. Этот механизм связан с величиной и формой половых органов самца и самки. Например, у животных с внутренним оплодотворением половой член самца должен соответствовать строению половых органов самки, с тем чтобы было возможно введение спермы. У растений, опыляемых насекомыми, каждый вид связан с определенным видом опылителей, и успех опыления зависит от величины, формы, окраски и запаха цветка и его способности привлекать насекомых именно данного вида.
Наконец, существует несовместимость гамет. Гаметы, продуцируемые организмом с тем, чтобы произошла сингамия, или слияние мужского и женского пронуклеусов с образованием диплоидного ядра зиготы, должны узнавать друг друга и обладать для этого специальными опознавательными признаками. Среди животных это наиболее ярко выражено у видов с наружным оплодотворением, которые выделяют гаметы в окружающую среду, обычно водную. Морские ежи и другие иглокожие, для которых характерно наружное оплодотворение, обладают опознавательными признаками, предотвращающими межвидовой обмен генами. Нельзя не испытать известного зловещего очарования, представив себе тот хаос, который мог возникнуть, если бы выброшенные в океан гаметы начали соединяться случайным образом. У растений несовместимость гамет чаще всего проявляется в неспособности пыльцевого зерна одного вида, попавшего на рыльце другого вида, прорасти в столбик, в результате чего мужской пронуклеус лишается возможности достигнуть яйцеклетки. Очевидно, этот же механизм предотвращает самооплодотворение у многих однодомных растений.
Ко второй крупной категории - постзиготическим изолирующим механизмам - относятся те механизмы, которые вступают в действие после того, как произошло слияние гамет. Первый из них - это летальность гибридов. Образование гибридов возможно, по крайней мере в лабораторных условиях, однако они гибнут на той или иной стадии развития. Гибель может произойти либо вскоре после оплодотворения, либо на довольно поздней стадии развития; она бывает обычно вызвана либо неспособностью отцовского генома выжить и/или функционировать в материнской цитоплазме, т.е. в цитоплазме яйцеклетки, либо несовместимостью отцовского и материнского геномов. Эти механизмы продемонстрировали Дени и Браше (Denis, Brachet) в своем исследовании причин летальности при скрещиваниях между двумя видами иглокожих - Paracentrotus lividus и Arbacia lixula. Яйца P. lividus можно оплодотворить спермой A. lixula, и они начинают развиваться, однако гибридный зародыш гибнет до гаструляции. Причиной прекращения развития может быть утрата отцовской ДНК дробящимся яйцом вследствие элиминации отцовских хромосом во время клеточных делений. Кроме того, в этих экспериментах не наблюдалось усиления синтеза РНК, которое обычно происходит при гаструляции или непосредственно перед ее началом. Летальность гибридов несколько иного рода была обнаружена при скрещиваниях между другой парой видов иглокожих - Dendraster excentricus и Strongylocentrotus purpuratus (Whiteley, Whiteley). В этом случае зародыши проходили через стадию гаструлы, но не достигали нормальной личиночной стадии. Возможно, что такое прекращение развития вызвано нарушением экспрессии отцовского генома, потому что при этом не происходит синтеза белков, специфичных для отцовского вида.
Другая форма постзиготической изоляции - это стерильность гибридов: межвидовые гибриды жизнеспособны, но не оставляют потомства. Классическим примером служит мул. Стерильность по вызывающим ее причинам делится на две основные категории - хромосомную и генную. Хромосомная стерильность часто возникает в результате неспособности хромосом отцовского и материнского происхождения к нормальной конъюгации и расхождению во время мейоза, что приводит к массивному «нерасхождению» при первом мейотическом делении. На рис. 3-5 изображено нормальное течение мейоза (слева) и случай нерасхождения (справа).
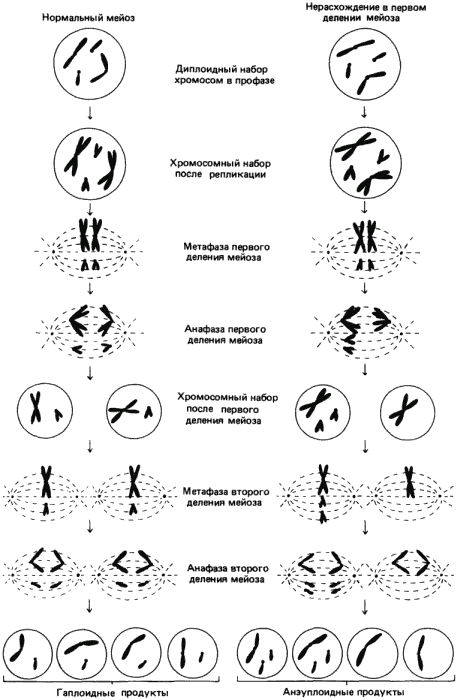
Рис. 3-5. Нормальный мейоз и аномальный мейоз, приводящий к нерасхождению хромосом. При нормальном мейозе все гаметы содержат равноценные гаплоидные наборы хромосом. При нерасхождении не происходит равномерного распределения хромосом по гаметам, так что гаметы содержат несбалансированные хромосомные наборы.
Как показывает этот гипотетический пример, аномальное распределение генетического материала при мейозе приводит к образованию анэуплоидных гамет, несбалансированных на хромосомном, а поэтому и на генном уровнях. Это происходит несмотря на то, что гибридный индивидуум во всех других отношениях совершенно нормален. Однако образуемые им анэуплоидные половые клетки неспособны соединяться ни друг с другом, ни с нормальными гаметами, с тем чтобы обеспечить нормальное развитие, а поэтому гибриды стерильны. Пример такой стерильности при скрещиваниях между двумя видами табака описали Клаузен и Гудспид (Clausen, Goodspeed). У Nicotiana tabacum диплоидное число хромосом равно 48, а у N. glutinosa оно равно 24. Эти два вида дают при скрещивании жизнеспособные гибриды с диплоидным числом хромосом, равным 36. Эти растения стерильны и не производят семян, потому что 12 хромосом N. glutinosa и 24 хромосомы N. tabacum, по-видимому, неспособны нормально конъюгировать и расходиться при мейозе.
Хромосомная стерильность другого типа может возникнуть при скрещивании двух близкородственных видов, обладающих различной хромосомной конституцией. Это легче всего наблюдать, если геном одного вида отличается от генома родственного вида по перестройке, состоящей в иной ассоциации плеч хромосом, т. е. если у одного вида произошла транслокация или слияние плеч хромосом. Гибрид от скрещивания между такими двумя видами продуцирует анэуплоидные гаметы вследствие аномального расщепления генетической информации, расположенной в них по-разному. У гибридного индивидуума, гетерозиготного по одному элементу транслокации, плодовитость будет понижена вдвое, а дальнейшие перестройки понизят ее даже еще больше. Аналогичное, хотя и менее выраженное, влияние хромосомные инверсии могут оказывать и оказывают на плодовитость внутривидовых гибридов. Следовательно, изменения числа хромосом и хромосомные перестройки могут блокировать обмен генетической информацией и создавать изоляцию, способствуя видообразованию.
Необходимо, однако, ясно понимать, что, устанавливая зависимость между изменениями генома (особенно это касается крупных изменений, различимых на глаз) и видообразованием, следует соблюдать осторожность и не путать причину и следствие. В некоторых случаях хромосомные изменения, несомненно, играют роль факторов, поддерживающих изоляцию, а не причин, вызывающих видообразование. Об этом свидетельствуют два примера. Один из них состоит в том, что, как мы увидим ниже, среди гавайских Drosophilidae часто происходит видообразование, не сопровождающееся хромосомными перестройками. Кроме того, среди этих же самых Drosophilidae встречаются многочисленные случаи полиморфизма по инверсиям, который поддерживается стабилизирующим отбором в популяциях, принадлежащих к одному виду, не приводя к кладогенезу. Полиморфизм по инверсиям можно лучше всего интерпретировать как существование блоков сцепленных генов, подвергающихся нормальному расщеплению, подобно сбалансированному полиморфизму в этой же популяции. Эти блоки, по-видимому, функционируют как супергены, обусловливающие гетерозис, потому что большая часть кроссоверов в пределах данной инверсии не попадает в ооцит, а элиминируется в полярных тельцах во время мейоза у самок. У самцов дрозофилы обычно не наблюдается кроссинговера при мейозе. Поэтому инверсии не снижают плодовитости. В этом отношении они отличаются от транслокаций. Хромосомные перестройки не всегда бывают связаны с видообразованием; и их присутствие необязательно свидетельствует о наличии процесса видообразования. При генной стерильности гибридные особи одного или обоих полов стерильны обычно вследствие аномального гаметогенеза. Нарушения возникают либо до, либо после мейоза. Пример этого в сочетании с интересным типом летальности гибридов наблюдается при скрещиваниях между двумя видами-двойниками - Drosophila melanogaster и D. simulans. Эти два вида морфологически идентичны, различаясь лишь по строению половых придатков самцов. В метафазе их кариотипы идентичны; при изучении их политенных хромосом обнаружена одна большая инверсия в 3-й хромосоме и пять или шесть мелких инверсий, разбросанных по остальному геному. Стертевант (Sturtevant) при скрещивании самок D. melanogaster с самцами D. simulans получал потомков только женского пола. При реципрокном скрещивании самок D. simulans с самцами D. melanogaster все потомство было мужского пола. В обоих случаях жизнеспособное потомство было полностью стерильным как при скрещиваниях между собой, так и при возвратных скрещиваниях с любым из родительских видов. Гонады у гибридов были мелкие и недоразвитые и не вырабатывали гамет.
Выявленный у этих гибридов тип летальности указывает, по мнению Стертеванта, на то, что для выживания гибридного потомства необходимо наличие Х-хромосомы D. simulans. Однако даже присутствия этой Х-хромосомы недостаточно для восстановления жизнеспособности, если в цитоплазме D. simulans присутствует также Х-хромосома D. melanogaster. Ключ к непосредственной причине этой летальности, а также подтверждение правильности гипотезы Стертеванта были получены в более поздних исследованиях этого явления. Дьюрика и Кридер (D. Durica, H. Krider) сумели показать, что в гибридных генотипах один из организаторов ядрышка (место синтеза рибосомной РНК) подавлен, что, очевидно, приводит к ослаблению синтеза белка. Большой интерес представляет доминантный аллель, недавно обнаруженный Такамурой и Ватанабе (Т. Takamura, Т. Watanabe) в природе у D. simulans; его присутствие спасает оба летальных класса при реципрокных межвидовых скрещиваниях. Мы вправе допустить, что этот аутосомный локус 2-й хромосомы ответствен за активацию или инактивацию рибосомных генов у межвидовых гибридов. Далее генетические исследования, проведенные несколько лет назад Мёллером и Понтекорво (H.J. Muller, G. Pontecorvo), позволяют считать, что число генов, контролирующих стерильность гибридов, невелико, но оно больше единицы. Хотя летальные взаимодействия, очевидно, детерминируются одним геном, находящимся во 2-й хромосоме, точное число, по-видимому, разбросанных по всему геному локусов, которые обусловливают стерильность жизнеспособного гибридного потомства, еще предстоит выяснить. Более четкая оценка генетической дифференцировки между двумя видами-двойниками дана в конце настоящей главы.
Последний тип постзиготической изоляции называют разрушением гибридов, или гибридным дисгенезом. При такой изоляции гибридные индивидуумы образуются, и они плодовиты; однако у потомков этих гибридных индивидуумов наблюдаются разнообразные нарушения развития: от летальности до пониженной жизнеспособности и стерильности. Примером этого явления служит скрещивание между Zea mays и ?. mexicana. Мангельсдорф (Mangelsdorf) получил от этого скрещивания гибридные растения, а затем скрещивал их с растениями Z. mays родительской линии. Потомки от этого скрещивания оказались высоко «мутабильными» и дали в свою очередь потомство с дефектами, затрагивавшими эндосперм, высоту стебля и другие признаки, от которых зависит мощность растений. Подобного рода эффект наблюдается не только у растений; недавно он был обнаружен и проанализирован у Drosophila melanogaster. В этом случае проявления дисгенеза можно видеть у гибридов от скрещиваний между мухами диких линий, выловленными в природе, и лабораторными линиями, которые были изолированы от диких линий в течение всего нескольких лет. Результаты дисгенеза проявляются в виде разрывов хромосом и их элиминации в процессе митотических клеточных делений у развивающихся личинок и в высокой частоте мутаций. Как показал генетический анализ этого процесса, он контролируется одним или несколькими генами и, возможно, связан с перемещением (транспозицией) небольших участков «кочующей» (мобильной) ДНК. Поэтому создается впечатление, что «разрушение» гибридов вызывается генами, влияющими на сохранение генома. В самом деле, возможно, что именно подобного рода механизм вызывает летальность гибридов, наблюдаемую в приведенном выше примере с иглокожими. Наконец, необходимо подчеркнуть, что репродуктивная изоляция редко бывает вызвана одной-единственной причиной; как в примере с гибридами между D. melanogaster и D. simulans, она может быть обусловлена несколькими факторами, блокирующими обмен генами. Некоторые из них могут быть пре-, а другие постзиготическими по своему действию. Конкретная комбинация факторов и природа механизма зависят от участвующих в гибридизации видов, от особенностей их поведения и физиологии размножения.
Механизмы изоляции генофондов сами по себе ничего не говорят о том, как первоначально возникла изоляция. Буш (Buch) обрисовал четыре возможных типа видообразования, которые если и имели место, то привели бы к кладогенезу. Эти четыре типа схематически представлены на рис. 3-6.
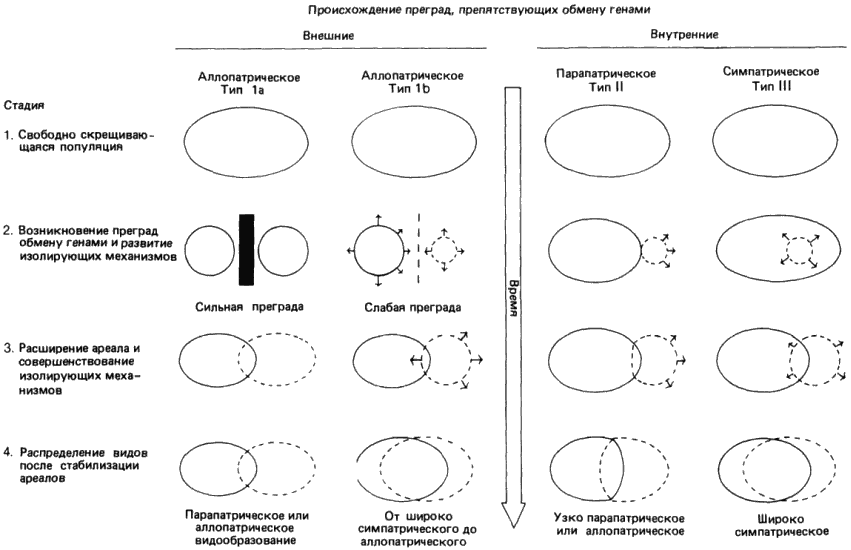
Рис. 3-6. Типы видообразования у животных (Bush, 1975).
Первый из них (тип 1а) обычно называют классическим, или аллопатрическим видообразованием. В исходном состоянии популяция представляет собой обширную панмиктическую (свободно скрещивающуюся) популяцию. Какое-либо изменение географии или климата разбивает эту большую популяцию на две отдельные части и при помощи той или иной физической преграды исключает возможность обмена генами между ними. Под действием отбора или просто в результате случайного дрейфа генетических элементов эти две некогда идентичные популяции превращаются в две генетически обособленные группы. Это изменение происходит не в результате одновременного изменения всех членов данной субпопуляции, а путем медленного замещения первоначальной генотипической конституции исходной популяции, происходящего за счет перераспределения генетического материала. Если физическая изоляция сохраняется на протяжении достаточно длительного времени, то возникает некоторый уровень генетической дивергенции, так что две новые популяции оказываются изолированными при помощи одного или нескольких внутренних изолирующих механизмов, которые мы уже обсуждали. Если на этой стадии устранить внешнюю преграду, то между такими прежде идентичными популяциями обмен генами происходить не будет. Таким образом, согласно определению, можно считать, что возникли два отдельных вида. Если (одновременно с репродуктивной изоляцией) у этих двух популяций появятся и другие признаки, в силу которых они станут занимать разные ниши, то они могли бы совместно обитать в одной и той же географической области (стать симпатрическими в результате миграции). В противном случае между ними может начаться конкуренция, которая приведет к вымиранию одного из двух видов.
Рис. 3-6 содержит еще одно уточнение, касающееся этого способа. Если репродуктивная изоляция несовершенна (особенно в тех случаях, когда возникает только постзиготическая изоляция), то возможен отбор, направленный на создание дополнительных преград. Существование одной лишь постзиготической изоляции привело бы к большой потере гамет для обеих популяций в любой точке их перекрывания, а любая особенность поведения, ведущая к презиготической изоляции, устранит эту потерю и будет подхвачена отбором. Следует указать, что видообразование такого типа протекает медленно.
Второй тип видообразования Буш назвал аллопатрическим видообразованием типа 1b. В этом случае процесс видообразования также начинается с разделения исходной панмиктической популяции на две географически изолированные группы. Разница состоит в том, что один из первоначальных изолятов мал по сравнению с другим. Малые размеры этого изолята могут серьезно повлиять на его исходный генофонд. Поскольку основатели новой маленькой популяции содержат лишь ограниченную выборку из генов обширной родительской популяции, по мере размножения этих основателей и освоения ими их новой среды может начаться формирование совершенно новой популяции. Если преграды остаются на месте в течение достаточно длительного времени, с тем чтобы могли накопиться существенные генетические изменения, то такая популяция превратится в самостоятельный вид и удаление преграды будет иметь те же самые последствия, что и при видообразовании 1а. Следует, однако, указать на основное отличие видообразования типа 1b от классического типа 1а: поскольку исходный изолят невелик, отбор и закрепление генетических различий могут происходить очень быстро, не требуя медленного замещения генов, необходимого в случае большой группы. Возможно, что быстрые и многочисленные разделения видов у гавайских Drosophilidae происходили именно при участии этого механизма «основателей»; кроме того, этот быстрый механизм и способ видообразования можно использовать для объяснения разрывов в палеонтологической летописи. И в самом деле, Темпльтон (Templeton), работавший с партеногенетическими линиями Drosophila mercatorum, продемонстрировал на одном экстремальном примере, в котором участвовал эффект «основателей», что событие, сходное с видообразованием, может произойти за одно поколение.
Описанные выше два типа видообразования эволюционные биологи принимают достаточно благосклонно; однако два других типа, представленные на рис. 3-6, вызывают больше возражений и, вероятно, возникают только при определенных обстоятельствах и у организмов, использующих специальные репродуктивные и экологические стратегии. Первый из них, обозначенный у Буша (рис. 3-6) как парапатрическое видообразование, или тип II, Уайт (М. White) называет также стасипатрическим. При этом способе видообразования изоляция обусловлена внутренними, а не внешними факторами. Первоначальный изолят, который очень мал по сравнению с исходной популяцией, образуется в результате изменения поведения особей-основателей. Событие, обусловливающее изоляцию, может выражаться в создании замкнутой системы скрещивания, или дема, как это бывает у грызунов, или же выбора нового хозяина, если речь идет о паразитическом организме. Благодаря такой изоляции обмен генами между группой основателей и родительским видом прекращается. В этом случае, снова из-за прекращения обмена генами, две новые популяции могут дивергировать. Обратите внимание, что в этой модели, поскольку первичное событие носит внутренний характер и приводит к репродуктивной изоляции, видообразование можно рассматривать как событие в геологическом масштабе времени почти мгновенное. Кроме того, ввиду того что начальная изоляция обусловлена внутренней причиной, родительская популяция и популяция основателей не обязательно должны быть разобщены географически.
И наконец, существует симпатрическое видообразование. При этом способе видообразования отделение, а поэтому и изоляция нового вида происходят целиком в пределах популяции родительского вида; т.е. процесс видообразования не сопровождается никакой географической изоляцией, которая бы допускала возникновение или поддержание репродуктивных изолирующих механизмов, обсуждавшихся выше. Для того чтобы произошло видообразование этого типа, изоляция должна возникнуть целиком и полностью и притом быстро в пределах ареала и местообитания родительского вида. Поэтому данный тип видообразования возможен только при наличии сугубо специфических условий и у организмов, в стратегии размножения которых имеется ряд необходимых для этого предпосылок. Интересно отметить, что этот способ, возможно, весьма обычен у растений. Почти мгновенное возникновение репродуктивной изоляции может происходить у растений в результате формирования тетраплоидов: два близких диплоидных растения изредка образуют вследствие нсрасхождсния при мейозе пыльцевые зерна и яйцеклетки с нередуцированным числом хромосом; гибриды между такими растениями бывают тетраплоидными (4n) в отличие от двух диплоидных (2n) родительских растений. Как указывает де Вет (de Wet), образование тетраплоидов чаще происходит в два этапа: сначала в результате слияния нормальной гаплоидной (n) и нередуцированной (2n) гамет получается триплоид (3n), а затем этот триплоид образует нередуцированную 3n гамету, которая при оплодотворении нормальной гаплоидной гаметой дает тетраплоид.
Процесс видообразования, происходящий за один этап, можно воспроизвести экспериментально, как это было сделано в скрещивании между редькой Raphanus saliva и капустой Brassica oleracea. У обоих растений число хромосом 2n = 18. Гибриды образуются, но они полностью стерильны. Девять хромосом, полученных от одного из родительских видов, не конъюгируют с девятью хромосомами, полученными от другого, и нормального расхождения также не происходит. Изредка, однако, от этих гибридных растений удается получить диплоидные пыльцу и яйцеклетки; это те случаи, когда ни одна из девяти хромосом не претерпела редукционного деления мейоза. У растения, получившегося в результате слияния 2n-пыльцевого зерна одного вида с 2n-яйцеклеткой другого вида, число хромосом 2n = 36. Это тетраплоид, содержащий 9 пар хромосом от R. saliva и 9 пар от В. oleracea. Он плодовит и воспроизводится путем самоопыления. Однако тетраплоидное растение дает плодовитое потомство только при самоопылении, но не при возвратных скрещиваниях с тем или другим из родительских видов. Благодаря своей новой хромосомной конституции оно репродуктивно изолировано от обоих родительских видов и представляет собой новый вид, хотя и полученный искусственно. В природе полиплоидия подобного рода может возникнуть в результате слияния гамет от одного и того же (аутополиплоидия) или от разных (аллополиплоидия) видов. В обоих случаях наблюдается та же картина, что и описанная выше: тетраплоид репродуктивно изолирован от обоих диплоидных родительских видов. Любое скрещивание между растениями 2n и 4n дает триплоидных (3n) потомков. Эти растения жизнеспособны, но они стерильны, потому что обычно продуцируют сильно несбалансированные или анэуплоидные гаметы. В нормальных случаях при мейозе хромосомы конъюгируют; конъюгация хромосом возможна при наличии четного числа хромосомных наборов. Если же число наборов нечетное, как у триплоидов (3n), то нормальная конъюгация и расхождение хромосом нарушаются. В некоторых гаметах хромосомы каждого данного типа бывают представлены в двух экземплярах, а хромосомы другого типа - только в одном. При соединении такой несбалансированной гаметы с нормальной гаплоидной гаметой получается зигота, также несбалансированная по своей хромосомной конституции. В большинстве случаев такое состояние летально. Только при скрещивании одного такого тетраплоида с другим тетраплоидом они дают плодовитое 4n потомство. Изоляция подобного типа возникла в группе видов Gilia transmontana. Это низкорослые однолетние травянистые растения, произрастающие в пустыне Мохаве, в западной части США. Как указывает Дэй (Day) и независимо от нее Грант (Grant), в эту группу входит 5 самостоятельных видов, которые обычно самоопыляются. Два из этих видов - G. transmontana и G. malior - тетраплоиды, тогда как три других - G. minor, G. clokeyi и G. aliquata - диплоиды. Все экспериментальные скрещивания между любыми двумя из этих пяти видов оказались стерильными. Поэтому, несмотря на морфологическое сходство между этими видами и на их симпатричность, они не скрещиваются между собой. На основании морфологического сходства Дэй пришла к выводу, что G. transmontana - тетраплоидное производное двух диплоидов, G. minor и G. clokeyi, тогда как также тетраплоидный вид G. malior произошел от G. minor и G. aliquata. Современное распространение этих видов в сочетании с их хромосомной конституцией можно рассматривать как непрямое доказательство их возможного симпатрического происхождения.
В связи с этим способом видообразования следует привлечь внимание еще к одной особенности растений. Многие растения однодомные (мужские и женские цветки находятся на одном растении) и самоопыляющиеся. Благодаря этой их черте полиплоидия может выступать в качестве мгновенно возникающего изолирующего механизма. В норме продуцирование диплоидных мужских и женских гамет - событие редкое. Их последующее слияние происходит еще реже. Поэтому вероятность возникновения двух тетраплоидных организмов, которые могли бы затем скрещиваться между собой и дать начало новому виду, ничтожна. Однако способность растений к самооплодотворению устраняет это затруднение. У животных, которые в большинстве своем двудомны, развитие изолирующих механизмов такого типа маловероятно. Кроме того, механизмы определения пола у животных, почти всегда связанные с хромосомами или генами, затрудняют возникновение полиплоидии. Любое изменение дозы хромосом или генов, затрагивающее гены или хромосомы, связанные с определением пола, приводит к нарушениям в определении пола и к стерильности, а поэтому у животных полиплоидия действительно встречается очень редко. Следует отметить, однако, что в тех случаях, когда репродуктивные затруднения преодолены, как при партеногенетическом размножении, полиплоидия у животных все же наблюдается.
Вероятно, наилучшее представление о многочисленности особых случаях видообразования, а следовательно, и эволюции у растений дает следующая краткая формулировка Гранта, указывающая на важную роль алло- и автополиплоидии у высших растений:
«Основные особенности растений повлияли не только на природу вида у растений, но оказали также глубокое воздействие на их макроэволюцию... в филогенезе растений неоднократно имело место образование анастомозов между линиями, которые прежде были обособленными. Если у животных видообразование обычного типа приводит к формированию филогенетического древа, то у растений оно нередко ведет к образованию филогенетической сети». Это утверждение не следует понимать в том смысле, что симпатрическое видообразование присуще только растениям и происходит только путем образования полиплоидов. Просто механизм, в основе которого лежит полиплоидия, - самый эффективный и быстрый. Рассматривалась также возможность симпатрического видообразования у животных при участии этологической изоляции. Буш (Bush) обратился к этому способу, чтобы объяснить видообразование у некоторых видов насекомых, паразитирующих на растениях. Изменение одного гена может повлиять на выбор хозяина этими насекомыми, а если всего лишь один индивидуум выбирает себе в качестве хозяина новое растение вместо прежнего, то это приведет к его изоляции от всей остальной видовой популяции.
Существуют разнообразные способы видообразования и всевозможные механизмы, при помощи которых у растений и животных может возникнуть репродуктивная изоляция. Но в этом участвуют еще два важных элемента. Первый из них - время, которое занимает данное «событие», а второй - степень генетической дивергенции, необходимая для того, чтобы возникла изоляция. Поскольку общеизвестно, что аллопатрическое видообразование поглощает много времени, мы не будем пытаться доказывать это, а постараемся найти минимальные оценки двух параметров - времени и генетической дивергенции, исходя из допущения, что любые значения, превышающие эти оценки, должны быть достаточны также для возникновения изоляции и видообразования.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
4.1. Типы взаимодействия
4.1. Типы взаимодействия Выделяют следующие типы взаимодействия между популяциями: «1) нейтрализм, при котором ассоциация двух популяций не сказывается ни на одной из них; 2) взаимное конкурентное подавление, при котором обе популяции активно подавляют друг друга; 3)
2.3. Вопросы макроэволюции и видообразования
2.3. Вопросы макроэволюции и видообразования Поскольку приспособительные изменения популяций (микроэволюция) разительно отличаются от картины разнообразия органического мира (макроэволюция), постоянно идет спор о наличии в макроэволюционных процессах особых факторов,
В поисках генов видообразования
В поисках генов видообразования Итак, постзиготическая изоляция возникает в результате фиксации несовместимых (конфликтующих) аллелей в разобщенных популяциях. Что же это за гены, изменения которых порождают такие конфликты? В принципе при скрещивании представителей
Цепная реакция видообразования
Цепная реакция видообразования Теоретически видообразование может быть самоускоряющимся процессом. Появление новых видов создает новые ниши, которые могут быть освоены следующим «поколением» новых видов. Например, появление нового вида травоядных может
6. ТИПЫ ДВИЖЕНИЙ
6. ТИПЫ ДВИЖЕНИЙ Движения человека очень разнообразны, однако всё это разнообразие можно свести к небольшому количеству основных типов активности: обеспечение позы и равновесия, локомоция и произвольные движения.Поддержание позы у человека обеспечивается теми же
Типы брачных отношений
Типы брачных отношений Основными типами брачных отношений в мире животных являются моногамия и полигамия.МоногамияПри моногамии животные образуют более или менее стойкие супружеские пары.Чаще всего моногамия встречается у незрелорождающихся млекопитающих, чьи
Типы взаимоотношений организмов
Типы взаимоотношений организмов Существует несколько классификаций типов взаимоотношений живых организмов в природе. Можно выделить основные варианты: конкуренция, протокооперация, комменсализм, мутуализм, паразитизм, хищничество, аменсализм.Конкуренция – это
Для видообразования нужны популяции, а не отдельные особи
Для видообразования нужны популяции, а не отдельные особи Селекция — вот путь, которым, по мнению большинства ученых, волки были превращены в собак. Однако эволюционирует не отдельная особь, а популяция, поэтому для видообразования нужно было, чтобы на протяжении
Дифференциальная выживаемость необходима для видообразования
Дифференциальная выживаемость необходима для видообразования Еще один принцип эволюционного видообразования путем искусственного или естественного отбора — дифференциальная выживаемость. При искусственном отборе это означает, что селекционер одним животным дает
Типы племенных кобелей
Типы племенных кобелей Племенных кобелей можно разделить на четыре типа.Первый тип — активный и опытный кобель, который не требует никаких предварительных церемоний и делает свою работу деловито и без суматохи. Кобеля такого типа нужно немного сдерживать, особенно с
Психологические типы А и Б
Психологические типы А и Б Человек учится всю жизнь: до трех дет – ходить и говорить, а потом всю оставшуюся жизнь – сидеть и молчать. Два основных типа поведения при стрессе: реакция «борьба или бегство» и «реакция затаивания» Типы стрессорной реакции человека и
Другие психологические типы
Другие психологические типы При рассмотрении склонности к поведению типа А или Б как основы психологического типа на первый план выходит такой компонент этого понятия, как стиль реагирования, т. е. особенности стрессорной реакции. Набор потребностей у носителей
Типы ритмов
Типы ритмов Золотистые водоросли демонстрируют суточный ритм, хотя их сутки и составляют 24,8 ч. Подобные ритмы называются циркадианными (от латинских слов circa — около и dies — день) или околосуточными. Цикл сна и бодрствования у человека, суточные колебания температуры
2.3. Основные типы пищеварения
2.3. Основные типы пищеварения По механизму действия ферментов на субстраты, по механизмам их поступления к месту функционирования, по взаимоотношениям пищеварительных процессов и клеточной границы и, наконец, по отношению к процессам транспорта можно выделить три