Возникновение сегментов
Возникновение сегментов
Сегментация зародыша дрозофилы в своей основе выглядит как ряд латеральных впячиваний зародышевой полоски, образующихся почти одновременно на стадии гаструляции. Несмотря на кажущийся мозаичный характер этого процесса, можно показать, что индивидуализация каждого сегмента происходит постепенно, начиная с дробления, до того как отдельные сегменты окончательно обособятся. Наложение на зародышей лигатур на стадии дробления, произведенное в нескольких лабораториях (Vogel; Herth, Sander; Schubiger, Wood), показало, что становление специфических особенностей каждого отдельного сегмента можно нарушить в разной степени, в зависимости от возраста дробящегося зародыша. Чем раньше накладывается лигатура, тем серьезнее нарушение. Полный набор сегментов детерминируется не раньше, чем будет достигнута стадия клеточной бластодермы. Во время дробления, по-видимому, должно происходить взаимодействие между различными участками зародыша, необходимое для возникновения нормальной сегментации. Возможная природа этого взаимодействия и генетическая регуляция процесса сегментации рассмотрены в ряде изящных морфогенетических и генетических исследований, проведенных Нюсслейн-Фольгардом и Вейсхаузом (Niisslein-Volhard, Weischaus).
Как уже отмечалось в гл. 7, Нюсслейн-Фольгард охарактеризовал два гена, наследуемые по материнскому типу, которые резко нарушают полярность зародыша. Мутации в этих двух локусах, bc (bicaudal) и dl (dorsal), приводят к тому, что гомозиготные мутантные самки продуцируют соответственно зародышей с двумя задними концами или с двумя дорсальными сторонами тела. Возможно, что эти два локуса содержат гены, продукты которых создают исходную позиционную информацию, детерминирующую основные передне-заднюю и дорсовентральную оси зародыша. Кодируют ли эти локусы саму информацию или же они необходимы для ее переработки - не известно. Однако, поскольку эти гены наследуются по материнскому типу, предполагается, что эта информация встраивается в яйцо во время оогенеза. На основе результатов экспериментов с наложением лигатуры можно думать, что окончательное становление сегментов происходит в результате дальнейшего уточнения информации об этих основных осях. Вместе с тем дело не ограничивается простым механическим распределением информации, поставляемой материнским организмом. Число сегментов, их местоположение, полярность и индивидуализация - все это должно быть определено (специфицировано). В одном из аспектов такой спецификации участвуют гомеозисные локусы, обсуждавшиеся в гл. 8. Эти локусы принимают участие в спецификации индивидуальных особенностей отдельных сегментов; они, по-видимому, действуют главным образом после оплодотворения и не проявляют материнского эффекта. Другие гены определяют число сегментов и их полярность. Недавно Нюсслейн-Фольгард и Вейсхауз обнаружили гены, детерминирующие все более детальное расчленение основных осей зародыша на определенное число сегментов. Подобно гомеозисным генам, определяющим индивидуальные особенности отдельных сегментов, эти гены экспрессируются в течение зародышевого развития. При помощи генетических и эмбриологических тестов была идентифицирована группа из 20 таких локусов, определяющих характер сегментации и особенности отдельных сегментов. Мутации в этих локусах не имеют материнского эффекта; они приводят к образованию фенотипов, у которых нарушено формирование сегментов, и являются летальными. Эти мутации не охарактеризованы полностью в генетическом и морфологическом отношении, поэтому неясно, все ли 20 генов специфически участвуют в сегментации или некоторые из них оказывают на этот процесс плейотропное воздействие. Однако 20 идентифицированных локусов, по-видимому, близки к общему числу таких генов в геноме дрозофилы.
Несмотря на предварительный характер анализа этих локусов, дефинитивные фенотипы мутантов дают некоторые указания о характере участия генов в становлении сегментации у зародыша. Более того, благодаря этим мутантам возникли интересные предположения об участии гомеозисных генов в вероятной интеграции процесса сегментации и в спецификации индивидуальных особенностей отдельных сегментов. Гены, определяющие сегментацию, можно разбить на основании создаваемых ими дефинитивных фенотипов на три широкие категории. Во-первых, это gap-локусы, приводящие к потере групп смежных сегментов; во-вторых, pair-rule-локусы, обусловливающие попарное слияние соседних сегментов, что приводит к уменьшению числа сегментов вдвое; и в-третьих, это локусы, изменяющие полярность сегментов; при этом зародыш содержит нормальное число сегментов, но каждый из них состоит из двух передних половинок, зеркально обращенных друг к другу.
Мутанты по двум из gap-локусов имеют фенотипы, представляющие интерес для обсуждаемой нами темы. Это мутации knirps и Rg (pbx) (Regulator of postbithorax). В результате мутации в локусе knirps развиваются зародыши с нормальным передним и задним концами тела, но с единственным большим брюшным сегментом там, где в норме должны находиться брюшные сегменты 1-7. Этот фенотип схематически изображен на рис. 9-8. Делеции локуса Rg (pbx) приводят как бы к реципрокному «выпадению» сегментов по отношению к выпадению, вызываемому делецией knirps. Вместо трех грудных и части гнатоцефалических сегментов образуется один большой сегмент, сходный с брюшным. Кроме того, седьмой и восьмой брюшные сегменты не разделяются. В обоих случаях из больших участков клеточной бластодермы и зародышевой полоски, которые в норме образуют несколько сегментов, формируется только по одному крупному структурному элементу. В широком смысле такое нарушение можно интерпретировать как результат какого-то дефекта в генетической программе клеток, находящихся в специфичных участках зародыша; этот дефект обусловливает неспособность клеток реагировать на информацию, поставляемую материнским организмом через яйцо. В пользу такого утверждения свидетельствует фенотип зародышей с делецией как локуса Rg (pbx), так и комплекса генов bithorax (BX-C). Делеция комплекса ВХ-С приводит к превращению брюшных сегментов в грудь. У зародышей с двойной делецией число сегментов такое же, как у зародышей Rg (pbx)- , но все эти сегменты грудные. Поэтому выпадение, наблюдаемое у зародышей Rg (pbx), приводит к нехватке некоторого участка тела у раннего зародыша и не зависит от наличия сегментов, обладающих особенностями грудных. Другими словами, ген Rg (pbx)+ определяет способность передней части зародыша к образованию сегментов и действует до индивидуализации сегментов или независимо от этого. В связи с этим последним соображением интересно отметить, что выпадающие у зародыша Rg (pbx) передние сегменты-это именно те сегменты, индивидуализация которых детерминируется комплексом ANT-C (Antennapedia). Поэтому правильное функционирование этого gap-локуса - необходимая предпосылка для экспрессии комплекса ANT-C.
Среди pair-rule-генов обнаружены мутанты двух типов. Примером одних служит локус prd (paired), примером других - локус ftz (fushi tarazu). Мутация prd приводит к «слиянию» задней части одного сегмента с передней частью другого в норме следующего за ним сегмента. Зародышевая полоска мутантного зародыша образует только половину нормального числа сегментов, каждый из которых вдвое шире нормального. Эти двойные сегменты схематически изображены на рис. 9-8; они состоят из средне- и заднегруди, первого и второго брюшного сегментов, третьего и четвертого брюшного и т.д. Возможно также слияние нижнегубного сегмента и переднегруди. Другой локус, ftz, входит в состав комплекса ANT-C (см. рис. 9-8), занимая на генетической карте место между гомеозисными локусами pb (proboscipedia) и Scr (Sex combs reduced). Так же как и локус prd, он приводит к «слиянию» сегментов, однако это слияние сдвинуто относительно того, которое вызывается локусом prd. Как видно на рис. 9-8, сливаются максиллярный и губной сегменты, первый и второй грудные, третий грудной и первый брюшной, второй и третий брюшные и т. д. Наблюдения над ранними зародышами показали, что это нарушение сегментации выявляется даже на начальных ее стадиях. Зародышевая полоска делится на вдвое меньшее число сегментов, чем в норме, причем все сегменты вдвое шире обычных. Исследование температурочувствительных аллелей локусов prd и ftz показало, что их температурочувствительные периоды (ТЧП) приходятся на очень раннюю стадию развития. Для аллеля ftz ТЧП наступает примерно через 2 ч после начала развития, что соответствует стадии клеточной бластодермы. Кончается ТЧП примерно спустя 4 ч после начала развития, прежде чем зародышевая полоска физически сегментируется. Таким образом, на стадии раннего зародыша происходит генетически регулируемое событие, необходимое для правильного осуществления сегментации.
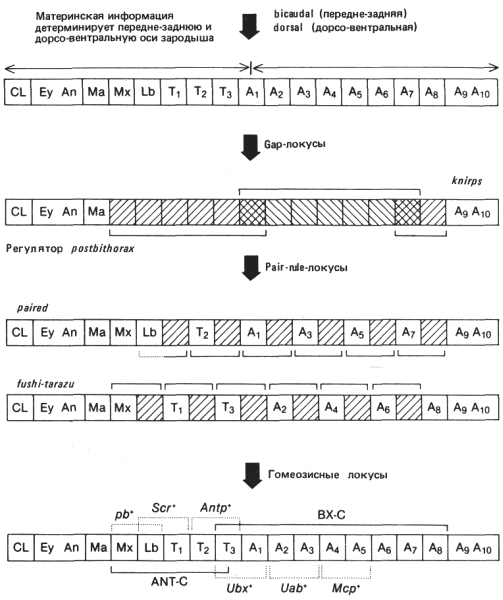
Рис. 9-8. Гипотетическая последовательность решений, принимаемых в зародыше дрозофилы на ранних стадиях развития для того, чтобы детерминировать индивидуализацию сегментов. Расшифровку буквенных обозначений см. рис. 8-9. Скобки над и под квадратиками указывают домены действия соответствующих генов. Штриховкой обозначены участки зародыша, в которых при мутациях gap и pair-rule нарушена нормальная сегментация. На нижней схеме указаны предполагаемые участки действия гомеозисных локусов; гены приведены в качестве примеров и не могут дать представления об огромном перечне известных генов, оказывающих влияние на характер сегментации. Данная модель описывает в упрощенном виде генетическую регуляцию сложного процесса, который в настоящее время интенсивно изучается.
Отсутствие этой генетической активности также показывает, что в какой-то момент детерминации характера сегментации зародышевая полоска расчленяется на ряд единиц шириной в два сегмента. Подобно локусу Rg (pbx), функция локуса ftz необходима для правильной экспрессии гомеозисных локусов. При попарном «слиянии» сегментов, наблюдаемом у зародышей ftz -, у широкого сегмента, образованного путем слияния двух соседних, проявляются только индивидуальные особенности переднего сегмента. Это наиболее ясно видно на примере слившихся третьего грудного и первого брюшного сегментов. У мутантных зародышей этот большой сегмент обладает только структурами, типичными для грудных сегментов; никаких признаков брюшных сегментов в нем не остается. Поскольку функция локуса bxd (bithoraxoid) заключается в спецификации индивидуальных особенностей первого брюшного сегмента, можно сделать вывод, что отсутствие функции ftz+ препятствует экспрессии локуса bxd. Однако активность ftz не связана с индивидуализацией сегментов. Зародыши, у которых отсутствуют локус ftz и комплекс ВХ-С, содержат половину нормального числа сегментов, но все это грудные сегменты. Этого фенотипа следовало ожидать, если локус ftz+, подобно Rg (pbx), вступает в действие раньше гомеозисных локусов, которые в свою очередь функционируют в пределах сегмента того типа, который установился путем спецификации, определяемой gap- и pair-rule-локусами.
В свете этого последнего момента следует также заметить, что слившиеся сегменты позволяют определить ассоциированные функции в пределах комплексов ANT-C и ВХ-С. Как схематически показано на рис. 9-8, областью третьего грудного - первого брюшного сегментов определяются границы территории, на которой действуют локусы bx+, pbx+ и bxd+. Все вместе эти гены не могут комплементировать доминантную мутацию Ubx. Сходные территории для действия других сайтов комплекса ВХ-С можно усмотреть в остальных задних спаренных сегментах. Если говорить о комплексе ANT-С, то мутации в локусе pb воздействуют на максиллярный и нижнегубной сегменты. Мутации Scr вызывают трансформации нижнегубного и первого грудного сегментов, а нуль-аллели гена Antp указывают, что для нормального развития второго и третьего грудных сегментов необходим аллель Antp+. Следовательно, эффекты генов, входящих в комплексы ВХ-С и ANT-С, показывают, что эти локусы действуют в области шириной в два метамера. Некоторые из этих областей можно определить по фенотипу ftz; другие выявляются по альтернативным типам слияния сегментов, наблюдаемым у мутантов prd. Подобно gap-локусам, pair-rule-локусы также определяют домены действия гомеозисных локусов, и именно эта их функция связывает между собой данные два вида генов, регулирующие сегментацию. Мутации по локусам, определяющим полярность сегментов, оставляют нормальными число сегментов и их индивидуальные особенности, но вызывают дупликации структур. Эти локусы, вероятно, наиболее близки по своим функциям к гомеозисным генам в том смысле, что они как бы расшифровывают позиционную информацию, которая в свою очередь определяет индивидуальные особенности, а не число метамерных элементов. На основе высказанных выше соображений можно построить связную и не чисто воображаемую картину способа становления сегментарной организации у зародышей насекомых.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
VI. Возникновение жизни на Земле
VI. Возникновение жизни на Земле Из опытов Спалланцани и Пастера мы уже знаем, что при высокой температуре жизнь прекращается. Большинство организмов погибает уже при 70–80 градусах тепла. Значит, для их жизни требуются определенные условия температуры. Требуются для
ВОЗНИКНОВЕНИЕ ЖИЗНИ
ВОЗНИКНОВЕНИЕ ЖИЗНИ Происхождение жизни, ее сущность — одна из наиболее трудных загадок науки, ибо жизнь — это самое сложное из известных нам явлений природы. Никто не видел и не наблюдал ее возникновения; более того, в природе не сохранилось никаких прямых или косвенных
ВОЗНИКНОВЕНИЕ КОЛОНИИ
ВОЗНИКНОВЕНИЕ КОЛОНИИ Образование отводка происходит обычно в определенные для каждого вида сроки. Вот как происходил начальный этап выделения нового муравейника у волосистого лесного муравья в 1967 г. в подмосковном ельнике (Солнечногорский лесокомбинат). В течение
Глава II. Возникновение нервных клеток и мозга
Глава II. Возникновение нервных клеток и мозга Причиной возникновения нервной системы стала низкая скорость получения информации о внешнем и внутреннем мире организма с донервной организацией. Его ткани состояли из клеток со сходной химической, электромагнитной и
§ 28. Возникновение отделов головного мозга
§ 28. Возникновение отделов головного мозга Ранний период истории возникновения предков позвоночных, до формирования хорошо структурированного скелета, довольно туманен. Если допустить, что предковые формы хордовых были мягкотелыми существами размером около 10–15 см,
§ 49. Возникновение мозга млекопитающих
§ 49. Возникновение мозга млекопитающих Небольшие рептилийные предки млекопитающих вышли из древесных завалов карбона с развитым обонянием, вестибулярным аппаратом, неважным зрением и ассоциативными центрами в среднем мозге. Эти существа начали загадочный
Возникновение и наследование модификаций на примере Metazoon
Возникновение и наследование модификаций на примере Metazoon Еще один важный теоретический вопрос, который можно рассмотреть на нашем примере с Metazoon, — это вопрос о возникновении адаптивных модификаций. Так называют способность организма более или менее осмысленно (то
Глава 2. Возникновение жизни: самозарождение и панспермия
Глава 2. Возникновение жизни: самозарождение и панспермия Трудно создать хорошую теорию, теория должна быть разумной, а факты не всегда таковы. Джордж У. Бидл, генетик, лауреат Нобелевской премии 1958 г. в области физиологии и медицины Физик Филипп Моррисон как-то заметил,
Возникновение филогенетической линии, ведущей к человеку
Возникновение филогенетической линии, ведущей к человеку Направление, ведущее к человеку, по мнению многих антропологов, берет свое начало от проконсула – крупной обезьяны, жившей около 20 млн лет назад в Африке, возможного родоначальника подсемейства дриопитеков.
Возникновение рода Homo
Возникновение рода Homo Поскольку эволюция гоминид происходила неравномерно, имела «мозаичный» характер, граница между древними представителями рода Homo и австралопитеками весьма размыта. Условными критериями отнесения к роду Homo принимаются объем мозга (не менее
Возникновение и эволюция человека современного типа
Возникновение и эволюция человека современного типа Происхождение современного человека – наиболее интригующая загадка антропогенеза. В антропологии все формы человека современного типа получили рабочее наименование «сапиенсы». Большинство их представителей были
Пролог Возникновение понимания
Пролог Возникновение понимания Галилей указал точку поворота, в которой научные усилия приняли новое направление, в которой ученые — анахронический, конечно, для того времени термин — поднялись со своих кресел, поставили под вопрос состоятельность прошлых попыток
Глава первая Эволюция Возникновение сложности
Глава первая Эволюция Возникновение сложности Без света эволюции ничто в биологии не имеет смысла. Феодосий Добжанский Великая идея: эволюция идет путем естественного отбораЖизнь столь совершенна, что, как долгое время считали, ее было необходимо сотворить особо. Ибо
4.2. Возникновение клеточной организации
4.2. Возникновение клеточной организации Учеными было доказано, что после испарения воды из реакционного объема в амфифильных липидоподобных и липидных молекулах формируются жидкокристаллические агрегаты, в которых молекулы расположены периодическими слоями, как в
9.3. Возникновение клеток
9.3. Возникновение клеток Предполагается, что этапом возникновения жизни на Земле следует считать период, когда сформировались простейшие клеточные системы, ставшие элементарной ячейкой живого. Сведения, касающиеся этой проблемы, освещены в обзорах, посвященных
9.3. Возникновение клеток
9.3. Возникновение клеток Предполагается, что этапом возникновения жизни на Земле следует считать период, когда сформировались простейшие клеточные системы, ставшие элементарной ячейкой живого. Сведения, касающиеся этой проблемы, освещены в обзорах, посвященных