Становление пространственной организации и спецификация местоположения
Становление пространственной организации и спецификация местоположения
Презумптивную область ноги, которую пересаживал Гаррисон, можно рассматривать как эмбриональное поле. В более общем смысле поле - это участок зародыша или группа клеток, в пределах которых может происходить регуляция. На примере поля конечности зародыша амфибии можно убедиться, что если удалить часть презумптивной конечности, то из оставшихся клеток тем не менее формируется нога со всеми ее обычными частями. Иными словами, оставшиеся клетки, если операция была произведена на достаточно ранней стадии развития, способны отреагировать на нехватку и заменить элементы, которые были удалены. Это установил Т. Морган (Т. Н. Morgan) еще до того, как он основал Американскую школу генетики. Он понимал, что такой процесс может происходить двумя основными способами: в первом случае недостающие структуры замещаются в результате клеточных делений, восполняющих удаленный материал; во втором случае регенерация поля происходит в отсутствие клеточного деления. Первый способ он назвал эпиморфозом, примером которого служит регенерация конечности у амфибий. Второй способ он назвал морфоллаксисом, наилучшим примером которого служит ранний зародыш в целом. У этих двух типов регуляции есть общая черта, состоящая в том, что клетки, образующие данное поле, способны определять и запоминать информацию о своем положении в пределах этого поля.
Для того чтобы объяснить регуляторное поведение клеток в пределах полей, Л. Вольперт (Lewis Wolpert) и его сотрудники сформулировали концепцию позиционной информации. Эта информация, сообщаемая клетке, содержит указание о ее местоположении по отношению к другим клеткам, и именно эта информация определяет план, в соответствии с которым развивается зародыш или поле. Чаще всего позиционная информация рассматривается как градиент какого-либо вещества (иногда называемого морфогеном), которое распределяется по полю так, что его концентрация постепенно изменяется от высокой к низкой.
Многие концепции позиционной информации возникли в результате исследований развития, начатых Сондерсом (Saunders) и его сотрудниками, а затем - работ Вольперта и его коллег на презумптивном зачатке крыла цыпленка. Почка крыла цыпленка развивается вначале так же, как почка конечности амфибии. По мере того как эта почка удаляется от главной оси тела, образуя лопастевидную конечность, из содержащейся в ней мезенхимы формируется хрящевой скелет конечности. Первыми появляются проксимальные элементы, а затем дистальные, т.е. пальцы. Дефинитивный скелет конечности изображен на рис. 9-2. На основании экспериментов с использованием меток из частиц угля были составлены карты зачатков элементов скелета. Было установлено, что по мере роста почки конечности, происходящего в проксимально-дистальном направлении, к ней добавляются все более и более дистальные структуры. Вначале появляется плечевая кость, затем лучевая и локтевая, позднее кости запястья и наконец фаланги пальцев. Эксперименты, проведенные Вольпертом и его сотрудниками, показали, что в процессе этого проксимально-дистального роста регуляция выражена слабо. Если они удаляли с дистального кончика почки конечности покрывающую ее эктодерму (которая не индуцирует морфогенез конечности) и часть лежащей под ней мезодермы, то дальнейший рост конечности прекращался. Если эти ткани удаляли в ранние сроки, то развивалась только плечевая кость. Если их удаляли немного позднее, то формировались плечевая, лучевая и локтевая кости, но ни запястье, ни пальцы не развивались. Такая последовательность во времени хорошо соответствовала карте зачатков, составленной по результатам экспериментов с применением меток. Эта карта изображена на рис. 9-2 (под схемой нормального крыла). Следует отметить, что удаление ткани из развивающейся почки производилось задолго до появления каких-либо признаков морфогенеза или клеточной дифференцировки. Следовательно, план строения конечности подвергался воздействию до того, как становились заметными явные признаки дифференцировки. Чтобы объяснить полученные результаты, Вольперт высказал мнение, что клетки мезодермы, лежащие под концевой верхушечной эктодермой, составляют то, чему он дал название прогрессивной зоны. Мезенхимные клетки в этой зоне не детерминированы, но в результате деления и роста на дистальном конце позади них, на проксимальной границе, остаются клетки, детерминированные к формированию специфических структур крыла; однако какие именно структуры будут формироваться, зависит от того момента времени, когда дочерние клетки покидают прогрессивную зону. Чем позднее данная клетка покидает эту зону, тем дистальнее формирующаяся структура. Поэтому судьба каждой клетки детерминируется в зависимости от времени, а не просто от ее местоположения в зачатке.
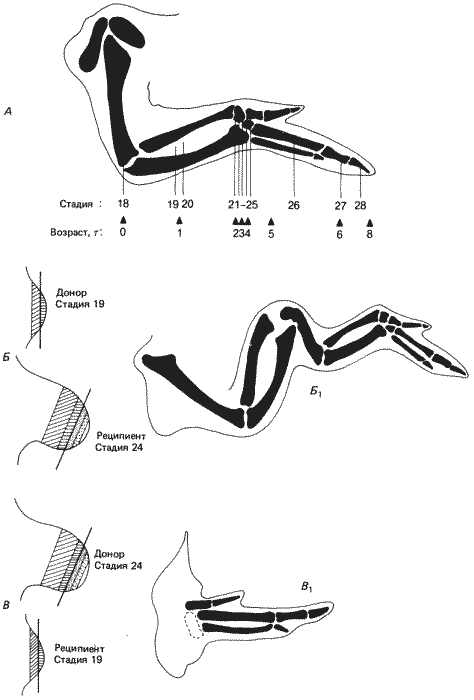
Рис. 9-2. Строение костных элементов крыла курицы (А) и результаты пересадок почек конечностей на разных стадиях развития (Б и В). Стадии развития на рис. А указывают сроки спецификации различных частей конечности: 18 - плечевая кость; 19 и 20 - лучевая и локтевая; 21-25 - область запястья; 26-28 - фаланги пальцев. Четыре выделенные на почках конечности области (Б и В) соответствуют зачаткам более проксимальных элементов конечности. Справа (Б1 и B1) показаны конечности, развивающиеся у реципиентов, которым были сделаны пересадки. У реципиента, находившегося на стадии 24, которому был пересажен кусочек более ранней почки конечности (стадия 19), образовалось крыло с тандемной дупликацией (Б1), тогда как при реципрокной комбинации образовалась конечность, в которой недостает проксимальных элементов (B1). (Wolpert, Lewis, Summerbel, 1975; с изменениями.)
Такой тип регуляции относится к классу эпиморфозов, потому что при этом необходимо клеточное деление. Для проверки этой гипотезы были произведены реципрокные пересадки предполагаемой прогрессивной зоны между «молодыми» и «старыми» почками конечности. Очень показательные результаты этих экспериментов схематически представлены на рис. 9-2. Если использовать в качестве донора почку конечности, у которой детерминированы только плечевая кость и проксимальные части лучевой и локтевой костей, а более «старую» почку, у которой детерминация достигла уровня запястья, - в качестве реципиента, то разовьется конечность с тандемно дуплицированными плечевой, лучевой и локтевой костями. При реципрокной пересадке наблюдаются нехватки элементов, которые дуплицировались в предыдущем эксперименте; образуется конечность, у которой запястье выступает прямо из боковой поверхности тела. Эти результаты полностью согласуются с гипотезой прогрессивной зоны и позволяют считать возможным, что положение определяется (специфицируется) не только в пространстве, но и во времени. Еще один момент, касающийся процесса спецификации, состоит в том, что позиционная информация не ограничивается только спецификацией судьбы данной клетки в чисто морфологическом аспекте. К такому выводу можно прийти, изучив область запястья крыла. Рассмотрение рис. 9-2 показывает, что развитие сравнительно маленького запястья занимает довольно много времени по сравнению с другими более крупными костями верхней конечности. Изучение относительной величины зачатков для каждого отдела верхней конечности показало, что изначально все они имеют одинаковые размеры; это означает, что миграция из прогрессивной зоны протекает равномерно во времени, различия же касаются динамики пролиферации разных отделов после спецификации. Запястье растет меньше, чем другие отделы крыла. Поэтому один из аспектов спецификации состоит, очевидно, в детерминации относительной скорости роста, а не только различной морфологии.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
2.1. Становление и основные положения синтетической теории эволюции
2.1. Становление и основные положения синтетической теории эволюции Эволюционизм возник как альтернатива учению о неизменности видов. Вопросы, связанные с возникновением и развитием жизни, прошли через всю интеллектуальную историю человечества. Количество литературы,
3.2. Становление этологии и вопросы эволюции поведения
3.2. Становление этологии и вопросы эволюции поведения Этология возникла в середине 1930-х гг. как наука, изучающая поведение животных в естественной среде обитания. Эта наука дала миру целую плеяду талантливых ученых, однако даже на таком ярком фоне выделяются имена ее
4.1. Типы социальной организации
4.1. Типы социальной организации В этологических исследованиях был обнаружен широкий диапазон различных видов социальной организации. У высших животных их социальные структуры и социальное поведение в процессе эволюции достигают исключительной степени сложности и
10.1. Становление взглядов на проблему когнитивных способностей животных
10.1. Становление взглядов на проблему когнитивных способностей животных Первые систематизированные данные по исследованию когнитивных способностей животных получил английский ученый, ученик Ч. Дарвина Дж. Романес (1848–1894). В 1883 г. выходит его книга «Интеллект животных».
11.1. Становление психологии
11.1. Становление психологии В дословном переводе «психология» означает «наука о душе». В научной терминологии вместо слова «душа» стали употреблять слово «психика», что, однако, не прибавило определенности этому понятию. Сложность, неопределенность, «неуловимость»
Становление локализации и пространственной организации
Становление локализации и пространственной организации Во время оогенеза происходит чрезвычайно активная транскрипция генов и накопление ооцитами мРНК. Накапливающиеся в ооцитах мРНК столь разнообразны по своим последовательностям, что, как только после начала
Эволюционные изменения в организации яиц хордовых
Эволюционные изменения в организации яиц хордовых Для Spiralia характерно развитие, при котором отдельные бластомеры на ранних стадиях дробления уже запрограммированы к дифференцировке в определенном направлении, не изменяющемся даже в том случае, если изолировать их от
Глава 9 Становление пространственной организации
Глава 9 Становление пространственной организации Пространственная организация и происхождение формы Главная проблема, стоящая перед биологами, занимающимися изучением развития, - объяснить механику процессов, в результате которых из одноклеточной зиготы образуется
Спецификация передне-задней полярности
Спецификация передне-задней полярности Можно представить себе, что спецификация передне-задней полярности развивающегося крыла устанавливается позиционной информацией примерно таким же образом, как устанавливается проксимально-дистальная ось. Однако, хотя
Становление пространственной организации и полярные координаты
Становление пространственной организации и полярные координаты Хадорн (Hadorn) и его ученики обнаружили, что если у личинки третьего возраста дрозофилы удалить один из имагинальных дисков и пересадить его путем инъекции новой личинке-реципиенту, то он будет
4.2. Возникновение клеточной организации
4.2. Возникновение клеточной организации Учеными было доказано, что после испарения воды из реакционного объема в амфифильных липидоподобных и липидных молекулах формируются жидкокристаллические агрегаты, в которых молекулы расположены периодическими слоями, как в
Становление ученого
Становление ученого Его жизнь прошла в беспрерывной борьбе и беспримерных дерзаниях. Десятилетиями длилась эта борьба. Трудно было поверить, что он обнаружит так много сил, найдет в себе столько решимости. Безудержная мысль Александра Васильевича Вишневского и на