Древесные саламандры и лягушки без головастиков. Гетерохрония и морфологическая адаптация
Древесные саламандры и лягушки без головастиков. Гетерохрония и морфологическая адаптация
Почему гетерохрония может служить таким обычным способом эволюции? Ответ на этот вопрос, по-видимому, заключен в характеристике, которую дал эволюции Ф. Жакоб (F. Jacob). По его словам, эволюция действует путем «перелицовки старого». В отличие от проектирования новых машин, создаваемых с учетом оптимальных технических возможностей, возможности эволюции ограничены историей. В данном случае история - это отдельные признаки или структуры, которые уже имеются у данного организма и могут быть модифицированы. Примером того, к каким это приводит результатам, служит прогрессивная эволюционная трансформация рудиментарных элементов нижней челюсти звероподобных рептилий в специализированные слуховые косточки внутреннего уха у млекопитающих. Морфогенетические процессы чрезвычайно сильно взаимодействуют между собой, и эти взаимодействия приводят к канализации развития. Согласно определению, данному в 1942 г. Уоддингтоном (Waddington), канализация - это забуферивание или гомеостаз путей развития, противодействующий извращениям развития, которые могут вызываться средовыми или генетическими возмущениями. П. Олберч (P. Alberch) высказал мнение, что кажущаяся направленность некоторых эволюционных линий отражает, возможно, ограничения, налагаемые теми эпигенетическими взаимодействиями, которые создают канализацию. Некоторые эволюционные изменения приводят к небольшим нарушениям существующих морфогенетических взаимодействий, тогда как другие теоретически возможные морфологические решения потребовали бы таких коренных изменений процессов развития, которые, в сущности, невозможны. Ввиду стабильности путей развития гетерохрония служит для эволюционных изменений своего рода путем наименьшего сопротивления. Относительные сроки различных процессов нередко можно изменять назависимо друг от друга, не вызывая резких нарушений канализации отдельных процессов, но тем не менее это может привести в конечном итоге к значительным изменениям морфологии.
Некоторые из наиболее ярких примеров эволюционной роли гетерохронических процессов получены в результате исследований, проведенных на амфибиях, в частности на тропических видах саламандр и лягушек, у которых в процессе эволюции возникли необычные отклонения в стратегии размножения. Большинство семейств хвостатых амфибий - обитатели умеренного пояса. Однако, как отмечает Уэйк (Wake), неотропические представители одной группы - трибы Bolitoglossini - составляют 40% всех ныне живущих видов саламандр. В один из родов этой группы, обширный и разнообразный род Bolitoglossa, входят не только типичные наземные формы, обитающие на возвышенных местах, но и единственные формы, успешно обосновавшиеся на низменностях тропических областей Центральной и Южной Америки.
Многие виды Bolitoglossa, будучи не наземными, что более обычно, а древесными формами, вполне приспособились к жизни на деревьях, и у них имеется ряд морфологических адаптации, в том числе очень цепкий хвост и модифицированные стопы, что дает им возможность удерживаться на нижних поверхностях влажных стеблей и листьев. Эволюция этих древесных видов сопровождалась уменьшением размеров тела взрослых особей и характерным комплексом изменений в строении стопы, показанных на рис. 6-3: наличие перепонок между пальцами и редукция или слияние костей. Концевые фаланги редуцированы, третья фаланга четвертого пальца утрачена, а некоторые из плюсневых костей слиты. Олберч сумел показать, что редуцированные размеры и своеобразное строение стопы у древесных форм прямо связаны с их способностью лазать по гладкой поверхности. Саламандры удерживаются при лазаньи главным образом благодаря поверхностному натяжению и присасыванию, обеспечиваемому строением стоп. Благодаря особенностям строения стопы у древесных видов Bolitoglossa они лучше удерживаются на ветвях при помощи присасывания, чем наземные виды, охватывающие ветви своими длинными пальцами. Уменьшение размеров тела, наблюдаемое обычно у древесных видов, также следует считать функциональной адаптацией, поскольку поверхностное натяжение более эффективно при небольших размерах.
П. Олберч и Дж. Олберч сравнивали древесный вид Bolitoglossa occidentalis с двумя наземными видами, В. rostrata и В. subpalmata. Как и ожидалось, древесный вид мельче двух других видов, у него имеются характерные модификации стопы и изменения в черепе: предлобные кости отсутствуют, окостенение черепа выражено в меньшей степени. По общим пропорциям взрослые В. occidentalis похожи на ювенильных особей В. rostratus и В. subpalmata: важно указать, что у ювенильных особей наземных видов на ногах имеются перепонки. Такое сходство позволяет предполагать, что в процессе эволюции древесных видов происходил отбор на небольшие размеры и что это достигалось путем прогенеза - урезания соматического развития.
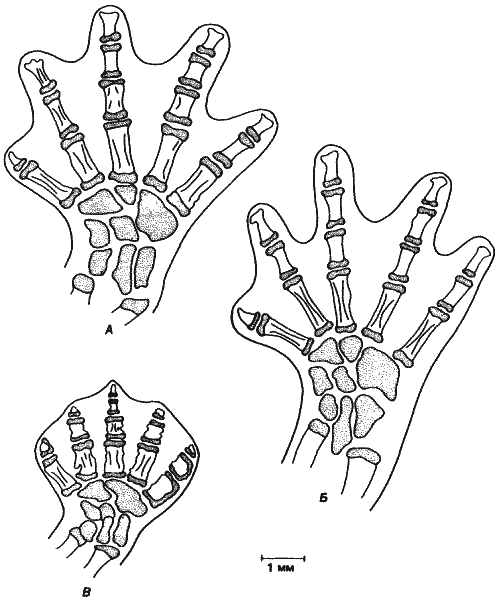
Рис. 6-3. Стопы наземных видов саламандры Boliloglossa (А и Б) и прогенетического древесного вида В. occidentals (В). Обратите внимание на относительно небольшие размеры стопы В. occidentalis, наличие у него перепонок и редукцию концевых фаланг (P. Alberch, J. Alberch, 1981).
Характер роста и аллометрические соотношения длины хвоста, массы тела, площади стопы и длины пальцев одинаковы у всех трех видов, как и следовало ожидать при простом прогенезе. В, occidentalis просто не вырастает до таких размеров, как другие виды. Рост у него прекращается на более ранней стадии, но при этом он достигает половозрелости. Возникновение педоморфной морфологии путем урезания развития представлено на рис. 6-4, где изображено изменение отношения длины пальцев к размерам перепонки в процессе развития разных видов Bolitoglossa. Вначале кинетика роста стопы из почки конечности практически одинакова у всех видов; однако пальцы растут быстрее, чем перепонки между ними. Но у В. occidentalis дифференциальный рост прекращается на той стадии, когда пальцы и перепонки оказываются одной длины. У двух других видов дифференциальный рост пальцев и межпальцевых участков продолжается, так что пальцы заметно выступают за пределы перепонок.
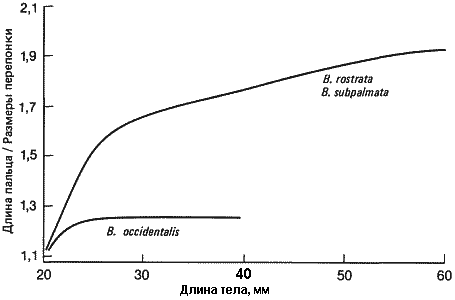
Рис. 6-4. Модели развития стопы у разных видов Bolitoglossa. У В. rostrata и В. subpalmata пальцы растут, удлиняясь и сильно выступая вперед и в стороны, тогда как у В. occidentalis пальцы никогда не выступают вперед, что приводит к развитию перепончатой лапы, наблюдаемой у взрослых особей (P. Alberch, J. Alberch, 1981).
При таком типе гетерохронии некоторые из ювенильных морфологических признаков древесных видов оказываются адаптивными, однако другие могут быть лишены адаптивного значения, будучи просто пассивными следствиями педоморфоза. Заключение о том, что рост Bolitoglossa occidentalis урезан, подтверждается существованием таких пассивных изменений, как зависимость между последовательностью появления костей стопы и черепа в процессе развития и отсутствие некоторых из этих костей у В. occidentalis. В черепе В. rostrata и В. subpalmata в последнюю очередь появляются предлобные кости, которые у В. occidentalis вообще не развиваются. Подобным же образом третья фаланга четвертого пальца в задней стопе В. rostrata и В. subpalmata развивается последней, а у В. occidentalis она совершенно отсутствует. Очевидно, развитие приостанавливается на стадии, непосредственно предшествующей появлению именно этих признаков.
Хотя уже глобальное изменение онтогенеза может открыть перед организмом новую адаптивную зону, для полной морфологической адаптации требуются последующие изменения в генетических системах, регулирующих отдельные аспекты морфогенеза. Р. Олберч и Дж. Олберч выявили такие дополнительные гетерохронии в морфогенезе стопы Bolitoglossa occidentalis. Развитие размеров и формы ноги разобщены, так что, когда длина животного (от кончика морды до анального отверстия) составляет 24 мм, изменение формы прекращается, но рост стопы продолжается, пока длина животного не достигнет 38 мм. В развитии стопы обнаружено разобщение и других процессов: скорость роста пястных костей повышена, а скорость роста некоторых фаланг понижена по сравнению с обычной для наземных видов скоростью. Это важный момент, поскольку он показывает, что гетерохрония не ограничена классическим глобальным типом, а может затрагивать любой отдельный процесс развития.
Подобно саламандрам, тропические лягушки проникли в некоторые адаптивные зоны, сильно отличающиеся от тех зон, в которых обитают виды, типичные для умеренного пояса. В тропиках популяция головастиков, развивающихся в водной среде, по-видимому, чрезвычайно сильно страдает от хищников и от конкуренции. Этот факт, а также наличие разнообразных влажных наземных микроместообитаний сыграли решающую роль в эволюции способов размножения, при которых лягушки не откладывают яйца просто в воду. В табл. 6-2 сопоставлены способы размножения у лягушек умеренного пояса США и у обитателей двух неотропических зон - в Панаме и Эквадоре. Почти все представители северных видов размножаются в открытых водоемах. Несколько видов, относящихся к тропическим группам, встречаются на юге США. Они дают представление о репродуктивном поведении, обычном для тропических фаун. Откладка яиц у поверхности воды или вблизи воды, так чтобы головастики попадали в пруд или реку после вылупления, сокращает время, в течение которого потомство подвергается превратностям жизни в воде. Эти адаптации возникли независимо в нескольких семействах лягушек.
Таблица 6-2. Разнообразие способов размножения лягушек умеренной и тропической фаун (Salthe, Duellman, 1973)
Биология размножения Доля всей фауны, % США Панама Эквадор Яйца и головастики в водоемах 90 20 37 Яйца расположены над водоемами; головастики в водоемах - 11 10 Яйца в пенистых гнездах; головастики в водоемах 1 11 8 Яйца и головастики в реках 1 11 4 Яйца расположены над рекой; головастики в реке - 11 4 Яйца на земле; головастики переносятся в воду - - 7 Яйца на земле; прямое развитие 6 30 21 Самка носит яйца на себе; прямое развитие - - 3 Не известна - - 6 Общее число видов в фауне 70 29 78С большей частью прогрессивной последовательности таких адаптации, устраняющих зависимость тропических лягушек от открытых водоемов, можно ознакомиться на примере одного рода Leptodactylus, рассмотренного Гейером (Heyer). В этот род входит пять групп видов. У групп видов Melanotus и Ocellatus наблюдается биология размножения, характерная для примитивных Leptodactylus, когда яйца откладываются в гнездо из пены, плавающее на воде. Представители эволюционно несколько более продвинутой группы Pentadactylus помещают свои пенные гнезда в рытвины, находящиеся поблизости от стоячих водоемов. Ливневые дожди разрушают эти гнезда, освобождая личинок и смывая их в водоем. Следующий шаг сделала группа Fuscus: самец роет норку и призывает к себе самку; яйца откладываются в гнездо из пены, помещаемое в норку. Развитие начинается в норке, но завершается в прудах, куда головастики смываются дождями. Полная независимость развития от водоемов достигнута в группе видов Marmoratus. Эти виды откладывают яйца в пенные гнезда в подземных камерах. Однако (в отличие от группы Fuscus) их яйца содержат достаточные запасы желтка, так что и развитие зародышей, и метаморфоз происходят в гнезде. Головастики все же образуются, но у них нет роговых челюстей с зубчиками и дыхалец, как у типичных водных головастиков.
Ряд эволюционных модификаций размножения, подобных описанным здесь на примере видов Leptodactylus, логически завершает прямое развитие с утратой личиночной стадии. Этого достигли представители нескольких семейств бесхвостых амфибий; новозеландская Liopelma (сем. Amphicoelidae), южноафриканская Arthroleptella (сем. Ranidae), африканские Breviceps и Ahydrophrene (сем. Brevicipitidae) и центральноамериканская Eleutherodactylus (сем. Bufonidae). Бесхвостые амфибии, у которых нет свободноживущей личиночной стадии, должны снабдить яйцо достаточным количеством желтка, чтобы обеспечить развитие ювенильной особи. Наиболее важная преадаптация, как считает Лутц (Lutz), состоит, по-видимому, в повышении содержания желтка в яйцах видов, цикл развития которых сходен с таковым некоторых видов Leptodactylus. Повышение содержания желтка, очевидно, служит еще одним примером генетически несложного изменения, создающего возможность для нового эволюционного направления. Интересно напомнить высказанное 50 лет назад предположение Нобля (Noble) о том, что увеличение содержания желтка в яйцах амфибий могло произойти внезапно, поскольку известны виды бесхвостых амфибий, продуцирующие как крупные, так и мелкие яйца.
Прямое развитие возникло в результате элиминации личиночных структур и ускорения развития признаков взрослого организма. В целом процесс этот не соответствует ни одному из классических определений гетерохронии, потому что при прямом развитии в результате коренным образом модифицированного онтогенеза образуется совершенно типичная лягушка. Наилучшее описание прямого развития дал Линн (Lynn) на примере тщательно исследованного им Eleutherodactylus nubicola. На рис. 6-5 развитие этой лягушки сопоставлено с развитием северо-американской Rana pipiens. В отличие от лягушек с хорошо выраженной водной личиночной стадией у Eleutherodactylus не развиваются органы боковой линии, кишечник никогда не бывает свернутым, а у Е. nubicola не развиваются и жабры. У некоторых других видов Eleutherodactylus жабры появляются, например у Е. portoricensis, которого изучал Гитлин (Gitlin). Утрата личиночных структур не была результатом некой глобальной гетерохронии, потому что одна очень существенная личиночная структура - хвост у представителей этого рода сохранилась, хотя и в модифицированном виде. Весьма вероятно, что утрата личиночных признаков отражает подавление индукционных систем, ответственных за появление отдельных личиночных структур. Одновременно происходила акцелерация появления некоторых дефинитивных структур. Наиболее ярко выраженная гетерохрония, проявляющаяся во внешних признаках, затрагивает зависимость между сроками развития конечностей и других структур. Сравнение относительных сроков развития конечностей и хвоста у Eleutherodactylus и Xenopus (рис. 6-6), у которой происходит типичный метаморфоз, позволяет выявить ряд различий в этих сроках. У Eleutherodactylus почки конечностей появляются очень рано - к концу замыкания нервной трубки. Рост конечностей у Eleutherodactylus начинается до начала дифференцировки главных отделов головного мозга и глаз, а у Xenopus и Rana - после нее. У Xenopus и Rana почки конечностей появляются только после начала быстрого роста хвоста, тогда как у Eleutherodactylus рост конечностей начинается до появления хвоста. Рост конечностей и хвоста протекает у Eleutherodactylus более или менее параллельно, а у Xenopus хвост растет быстрее, чем конечности, в росте которых происходит скачок лишь во время метаморфоза. У обоих видов конечности достигают полной длины, свойственной ювенильной стадии, после начала резорбции хвоста.
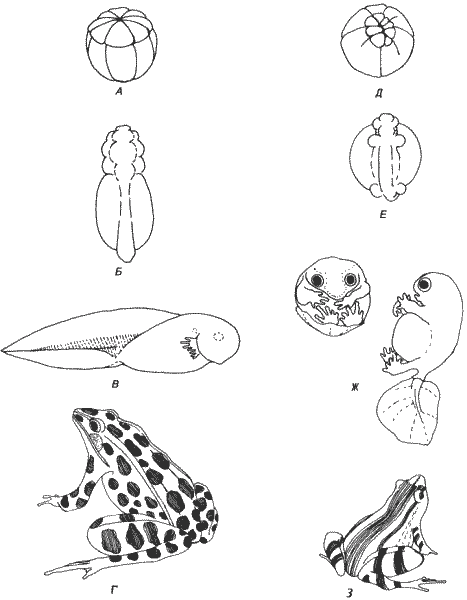
Рис. 6-5. Сопоставление развития, сопровождающегося метаморфозом, и прямого развития у лягушек Rana pipiens (А-Г) и Eleutherodactylus nubicola. А. Дробление яйца R. pipiens. Б. Нейрула. В. Головастик с жабрами и хвостом. Г. Взрослая особь. Д. Дробление яйца Е. nubicola. E. Нейрула с преждевременными почками конечностей и хвостовой почкой. Ж. Готовый вылупиться лягушонок, у которого хвост модифицирован в орган дыхания. З. Взрослая особь (эмбриональные стадии по Hamburger, 1960, и Lynn, 1942; взрослые особи - с натуры).
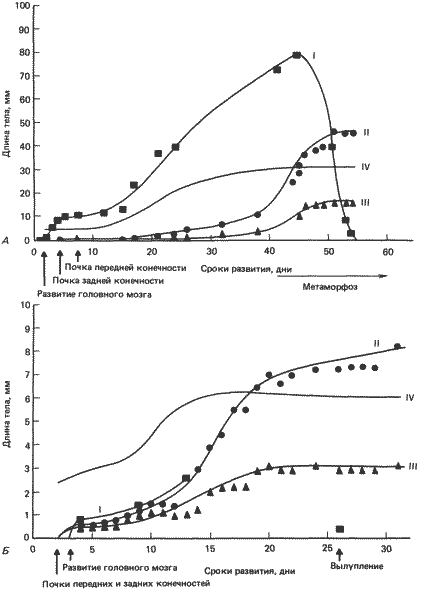
Рис. 6-6. Относительные сроки различных событий в процессе развития Xenopus и Eleutherodactylus. А. У Xenopus - лягушки, проходящей в процессе развития через стадию головастика и метаморфоз, - хвост появляется задолго до конечностей. Развитие головного мозга также успевает достаточно продвинуться до того, как начинается рост конечностей. У головастика конечности растут очень медленно. За этой фазой медленного роста конечностей следует взрыв их роста во время метаморфоза, причем хвост в это время быстро рассасывается. Б. У Eleutherodactylus - лягушки с прямым развитием - рост конечностей начинается до появления хвоста и до развития головного мозга. За исключением хвоста, рассасывающегося незадолго до вылупления, все остальные признаки достигают дефинитивного состояния путем прямого развития, и метаморфоза не происходит. I - хвост; II - задние конечности; III - передние конечности; IV - длина тела. (По данным Neiuwkoop, Faber, 1956, и с рисунка и по данным Lynn, 1942.)
Ускорение развития почек конечностей не обязательно отражается на других признаках. Так, например, Линн сравнивал последовательность и сроки окостенения черепа у Eleutherodactylus с таковыми у Rana temporaria, которую изучал Эрдман (Erdmann). Развитие черепа у Eleutherodactylus отличается от его развития у Rana одним очень заметным признаком - полным отсутствием у Eleutherodactylus супраростральных и инфраростральных хрящей, которые поддерживают ротовое отверстие головастиков Rana, самостоятельно питающихся до метаморфоза. У Rana до начала метаморфоза окостеневают только пять хрящей; во время метаморфоза окостеневают другие хрящи; несколько последних хрящей окостеневают после окончания метаморфоза. Порядок окостенения у Eleutherodactylus позволяет выявить несколько гетерохронических изменений по сравнению с Rana. В период примерно между 16-м и 19-м днями развития окостеневают первые хрящи. Появление угловой и чешуйчатой костей ускоренное по сравнению с их появлением у Rana. Вслед за этим происходит еще ряд окостенений, так что за четыре дня до вылупления окостенение черепа у Eleutherodactylus находится примерно на том же уровне, что у Rana при метаморфозе. Однако появление некоторых костей запаздывает. Septomaxillare, переднеушная и подбородочная кости, появляющиеся у Rana при метаморфозе, а также кости, появляющиеся у Rana вскоре после метаморфоза, у Eleutherodactylus спустя 10 дней после вылупления еще отсутствуют.
Если считать рост конечностей и резорбцию хвоста морфологическими показателями метаморфоза у Xenopus и Rana, то следует считать, что зародыш Eleutherodactylus достигает стадии, соответствующей метаморфозу, за несколько дней до вылупления. Однако к этому времени он отстает от метаморфизирующих личинок Rana не только по степени окостенения черепа, но и по развитию некоторых других элементов скелета. Например, Линн указывает, что, если у Rana во время метаморфоза седалищная кость, грудина и надгрудинник уже имеются, у Eleutherodactylus к моменту вылупления окостенение соответствующих хрящей не начиналось. Наиболее резко выраженное ускорение наблюдается у Eleutherodactylus в развитии дуг аорты. У зародышей позвоночных образуется шесть пар артериальных дуг, или дуг аорты, в соответствии с шестью парами глоточных, или жаберных, мешков. У челюстноротых рыб и высших позвоночных первая пара дуг во взрослом состоянии никогда не сохраняется полностью; у некоторых рыб сохраняется вторая пара, а у большинства - дуги III-VI. У четвероногих дуги I и II исчезают во время развития, так же как и дуга V, сохраняющаяся только у хвостатых амфибий. Дуги III, IV и VI образуют соответственно каротидную дугу, системную дугу и легочную артерию. Судьба артериальных дуг у бесхвостых амфибий показана на рис. 6-7, А-В, взятом из работы Милларда (Millard) no Xenopus laevis. У этого вида первая артериальная дуга появляется на ранней стадии личиночного развития, а вскоре после нее одна за другой появляются третья и четвертая дуги. Вторая артериальная дуга появляется лишь в виде зачатка. Первая и вторая дуги начинают дегенерировать еще до появления шестой дуги. Дуги III-VI снабжают кровью жабры. На более поздних стадиях развития исчезает пятая дуга, так что у взрослого животного сохраняются только дуги III, IV и VI.
На рис. 6-7 сопоставляется развитие артериальных дуг у Eleutherodactylus и Xenopus. Как и у Xenopus, артериальные дуги появляются у Eleutherodactylus на относительно ранних стадиях развития, но происходит это совершенно иначе. Первая и вторая дуги не появляются вовсе. Третья дуга появляется первой, за ней следует четвертая, а затем шестая. Таким образом, у этой лягушки ускоренным способом непосредственно образуется кровеносная система взрослого организма на ранних стадиях развития зародыша, у которого никогда не развиваются жабры и которому поэтому не нужна кровеносная система, связанная с жабрами. Особенно интересно, что дуги I, II и V не появляются у Eleutherodactylus даже в качестве временных структур. Для всех высших позвоночных - рептилий, птиц и млекопитающих - характерно прямое развитие и отсутствие жабер, тем не менее у зародышей рептилий и птиц появляются все шесть артериальных дуг, а у зародышей млекопитающих - все дуги, кроме пятой. Как и у Xenopus, у высших позвоночных во взрослом состоянии сохраняются только дуги III, IV и VI. Временное появление артериальных дуг у рептилий, птиц и млекопитающих позволяет предполагать, что они выполняют какую-то функцию в развитии, возможно, в качестве элементов каскадных индукционных взаимодействий, необходимых для появления каких-то других структур. У Eleutherodactylus эта функция, по-видимому, отпала.
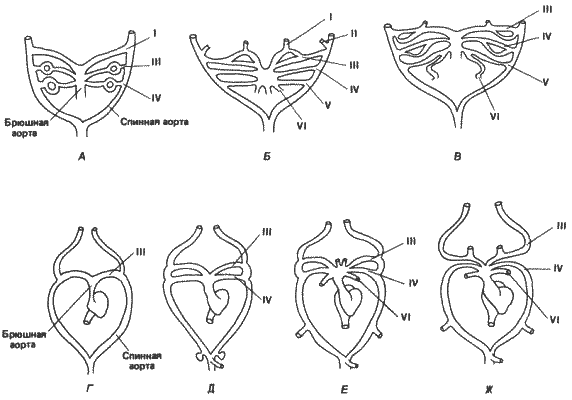
Рис. 6-7. Развитие артериальных дуг у Xenopus и у Eleutherodactylus. У Xenopus (А-В) появляются все 6 пар артериальных дуг, хотя дуги I и II сохраняются у нее очень недолго. Петли в дугах у Xenopus схематически изображают кровеносную систему жабр. Дефинитивная форма сердца и артериальных дуг после метаморфоза у Xenopus сходна с изображенными на схеме Ж. У зародышей Eleutherodactylus (Г-Ж) дуги I и II не появляются вовсе. Дефинитивное состояние достигается сразу в результате развития дуг III, IV и VI. Жабры совершенно не образуются (Millard, 1945; Linn, 1942).
Прямое развитие, наблюдаемое у Eleutherodactylus, иллюстрирует эволюционный переход к наземному образу жизни, независимый от того пути, по которому следовали в палеозое предки амниот. Однако представляется вероятным, что переход от размножения с прохождением через личиночную стадию, обитающую в воде, к размножению на суше у предков рептилий мог происходить таким же образом - путем ускорения развития важнейших признаков взрослой стадии. Гетерохрония при этом привела не к иной дефинитивной морфологии, а к иному типу онтогенеза, который в сочетании с эволюцией яйца амниот сделал возможной всю последующую эволюцию наземных позвоночных.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Лягушки и жабы
Лягушки и жабы Единственные земноводные, часто попадающиеся на глаза путешественнику в экваториальных местностях, – это лягушки и жабы, в особенности миловидные древесные лягушки. В начале дождливого времени года, когда высохшие лужи и рвы наполняются водой, лягушки
Полезная адаптация?
Полезная адаптация? Канадские психологи Ара Норензаян и Азим Шариф из Университета Британской Колумбии наряду с другими экспертами полагают, что религиозность вполне могла развиться как полезная адаптация, способствующая сплоченности коллективов (Norenzayan, Shariff, 2008).
Крысы и лесные лягушки
Крысы и лесные лягушки Интересный факт, который трудно укладывается в голове: оказывается, что если весь род человеческий, населяющий землю, поставить плотно сомкнутым строем в затылок друг другу, то он займет всего половину острова Уайт[8]. Как властелины земли, мы
Рассказ Саламандры
Рассказ Саламандры Названия – угроза эволюционной истории. Не секрет, что палеонтология – спорный предмет, где имеет место даже некоторая личная вражда. Напечатаны, по крайней мере, восемь книг под названием «Кости раздора». И если Вы посмотрите на то, из-за чего ссорятся
Если амфибия машет скалкой: проявления ревности самки саламандры
Если амфибия машет скалкой: проявления ревности самки саламандры Лесные саламандры известны не только своей страстностью. Они вполне сносно умеют карабкаться, прыгать и плавать. Однако проявления чрезмерного темперамента с их стороны невероятны, так как у саламандр
6. АДАПТАЦИЯ СЕНСОРНОЙ СИСТЕМЫ
6. АДАПТАЦИЯ СЕНСОРНОЙ СИСТЕМЫ Сенсорная система обладает способностью приспосабливать свои свойства к условиям среды и потребностям организма. Сенсорная адаптация – это общее свойство сенсорных систем, заключающееся в приспособлении к длительно действующему
2.11. Зрительная адаптация
2.11. Зрительная адаптация При переходе от темноты к свету наступает временное ослепление, а затем чувствительность глаза постепенно снижается. Это приспособление зрительной системы к условиям яркой освещённости называется световой адаптацией. Обратное явление
Тупайи: древесные землеройки в состоянии стресса
Тупайи: древесные землеройки в состоянии стресса Мы с знаем, что чрезмерный стресс вредит здоровью. Многочисленные тяжелые заболевания — рак, мигрень, артрит и дерматит, а также депрессии и психозы связаны именно с ним. Мы также знаем, что может подвергнуть нас
Гетерохрония
Гетерохрония Первую классификацию явлений гетерохронии предложил Г. де Бер, выделив 8 разных типов (DeBeer G., 1958). Затем различные авторы давали другие классификации и свои термины, что создавало терминологическую путаницу (Gould S., 1977).В самом общем виде все варианты
3.3.1. Адаптация растений к поддержанию водного баланса
3.3.1. Адаптация растений к поддержанию водного баланса Низшие наземные растения из влажного субстрата поглощают воду погруженными в него частями таллома, а влагу дождя, росы и тумана – всей поверхностью. В максимально набухшем состоянии лишайники содержат в 20–30 раз
Глава 3 Морфологическая и молекулярная эволюция
Глава 3 Морфологическая и молекулярная эволюция Я думаю, что отец наш небесный выдумал человека, потому что разочаровался в обезьяне. Марк
Гетерохрония - классический механизм эволюционных изменений
Гетерохрония - классический механизм эволюционных изменений Попытки найти механизмы эволюции, связанные своими корнями с онтогенезом, сосредоточены главным образом на гетерохронии - изменениях относительных сроков процессов развития. Геккель подчеркивал зависимость
Морфологическая и молекулярная рекапитуляция. «Треснувшее зеркало»
Морфологическая и молекулярная рекапитуляция. «Треснувшее зеркало» Джордж Уолд (G. Wald) начинает свое обсуждение молекулярной рекапитуляции со следующего утверждения: «Живые организмы представляют собой сильно увеличенные выражения тех молекул, из которых они
Квохтающие лягушки
Квохтающие лягушки Цель самца — соблазнение: он манипулирует самкой, чтобы заставить ее поддаться его чарам. Эволюционное давление заставляет его совершенствовать брачную демонстрацию, которая располагает к нему и сексуально возбуждает самку.В случае со скорпионами
Рассказ Саламандры
Рассказ Саламандры Названия – вечный камень преткновения в науке об эволюции. Ни для кого не секрет, что палеонтология противоречива, и в этой отрасли науки нередко вспыхивает вражда. (Есть по крайней мере восемь книг, озаглавленных “Кости раздора”.) Чаще всего
Наследие рыб и головастиков: икота
Наследие рыб и головастиков: икота Икота — неприятность, восходящая корнями к истории, роднящей нас с рыбами и головастиками.Если что-то и может нас в связи с этим утешить, так это то, что наше несчастье разделяют с нами и многие другие млекопитающие. У кошек можно