О зубах курицы и перьях ящерицы
О зубах курицы и перьях ящерицы
Если эволюционные изменения в самом деле возникли в результате диссоциации индукционных событий, то можно ожидать, что остатки прежних взаимосвязей должны сохраняться, что регуляторные гены, не выявляющиеся таким образом, как прежде, все еще присутствуют. Подобное ожидание вполне обоснованно, как показывает рис. 5-6, на котором О. Мангольд (Mangold) схематически изобразил некоторые из основных индукционных событий в голове и туловище зародышей амфибий. В этой схеме подчеркивается общее заключение, уже выведенное из работ Джекобсона, о сложности индукционных взаимодействий, которые, как правило, принадлежат к типу переплетающихся или каскадных взаимодействий. Применительно к эволюции это означает, что индукционные взаимодействия устойчивы к существенным модификациям и что в случае возникновения достаточно крупных изменений они обычно не выходят за рамки характерных для данной группы. Коренные изменения, с которыми сопряжено возникновение некоторых новых групп, таких как членистоногие (см. гл. 4), иногда имеют место, но это случается редко. Другое следствие интегрированности морфогенетических путей, о котором часто упоминают, состоит в том, что эволюционные изменения, относящиеся к поздним стадиям развития, воспринимаются легче, чем ранние изменения, просто потому, что поздние изменения требуют меньше соответствующих изменений в затрагиваемых ими каскадных процессах. Вряд ли, однако, можно считать, что изменения в онтогенезе возникают исключительно таким образом; если бы это был единственный способ, то что-то очень близкое к геккелевской рекапитуляции действительно играло бы универсальную роль. На самом же деле изменения возникают на всех стадиях развития, так что хотя картины взаимодействий, наблюдаемые у амфибий, и можно считать своего рода архетипом для развития позвоночных, среди последних нельзя найти двух видов, развитие которых протекало бы совершенно одинаково, пусть даже на самых ранних стадиях. Раннее сходство и все возрастающее в дальнейшем обособление зародышей родственных организмов, о которых говорится в законах Бэра, не следует понимать чересчур буквально. Развивающиеся млекопитающие ни на какой стадии развития не бывают идентичны зародышам рыбы или ящерицы: по своим геномам и характеру развития они очень далеко ушли от своих предков. Но тем не менее некое исконное сходство между ними сохранилось. А произошло следующее: множественные индукционные взаимодействия, описанные на примерах индукции хрусталика, носа и уха, создали достаточно высокий уровень морфогенетического гомеостаза, так что изменения могут восприниматься даже на ранних стадиях общего процесса развития, не нарушая его интегрированности.
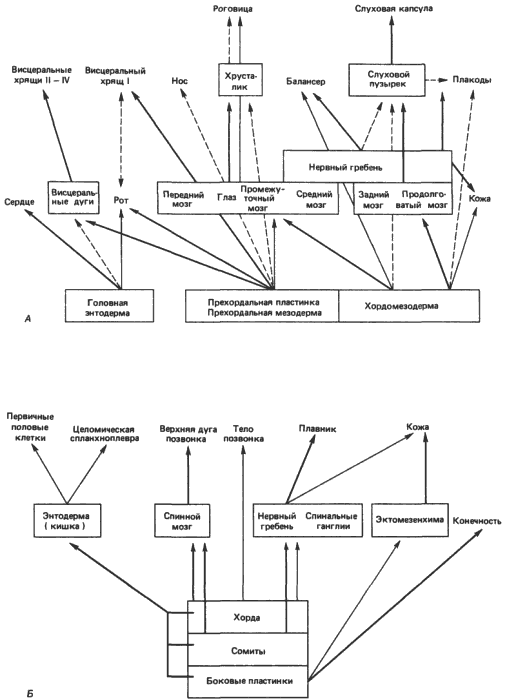
Рис. 5-6. Индукционные взаимодействия и каскадные процессы в развитии амфибий. А. Индукционные процессы в развитии головы. Б. Индукционные процессы в развитии туловища. Толщина стрелок отражает относительную интенсивность индукции (Mangold, 1961).
Существование общего консервативного плана развития позволяет считать, что эмбриональные структуры могут сохраняться даже после того, как они утратили свою прежнюю основную функцию, потому что они все еще служат связующими звеньями в каскаде морфогенетических взаимодействий, присущих прежней функции. Таким же образом могут сохраняться и регуляторные системы. Доказательством сохранения прежних регуляторных генов могло бы служить воскрешение отвергнутого в прошлом пути развития при нарушении существующего типа развития в результате мутации или экспериментального воздействия. Известны примеры того и другого. Восстановление старых типов развития в результате мутаций подробно рассматривается в гл. 8 и 9. Экспериментальный анализ трех примеров эволюционных изменений позволил обнаружить модификации в системах взаимодействия между тканями, при которых на прежние механизмы регуляции налагались бы новые, но без утраты генетической информации, имевшейся у предков. Два из этих примеров относятся к модификациям, имевшим место в эволюции птиц; третий касается изменений в покровных тканях рептилий, птиц и млекопитающих.
Археоптерикс издавна известен как «недостающее звено», поскольку он обладал такими птичьими признаками, как перья в ни с чем несообразном сочетании со скелетом, характерным для рептилий, а точнее - для динозавров, вплоть до длинного хвоста и зубов. Многие изменения, возникшие в процессе эволюции у птиц, привели к повышению эффективности полета путем снижения массы тела; это сопровождалось утратой некоторых характерных признаков археоптерикса, в том числе зубов, когтей на передних конечностях и длинного рептильного хвоста, а также редукцией или слиянием некоторых костей конечностей. Последние зубатые птицы относятся к позднему мелу, т. е. прошло достаточно много времени, чтобы геномы птиц могли освободиться от генетической информации, необходимой для морфогенеза зубов. На самом же деле этого не случилось, и у курицы можно действительно вызвать образование зубов.
Для нормального процесса развития зубов, как это указывается в обзорах Теслеффа (Thesleff) и Дьюкар (Deuchar), необходимы реципрокные индукционные взаимодействия между оральным эпителием и лежащей под ним оральной мезенхимой. Последовательность происходящих при этом событий схематически показана на рис. 5-7. Оральная мезенхима индуцирует в лежащем над ней эпителии развитие эмалевого органа; этот орган в свою очередь инвагинирует в мезенхиму и индуцирует ее дифференцировку в одонтобласты, которые секретируют дентин. В отсутствие эмалевого органа оральная мезенхима дает начало только губчатой костной ткани. В развитии зубов участвует еще один индукционный процесс: эпителиальные клетки внутренней части эмалевого органа реагируют на присутствие мезенхимальньгх одонтобластов, дифференцируясь в амелобласты, которые секретируют зубную эмаль.
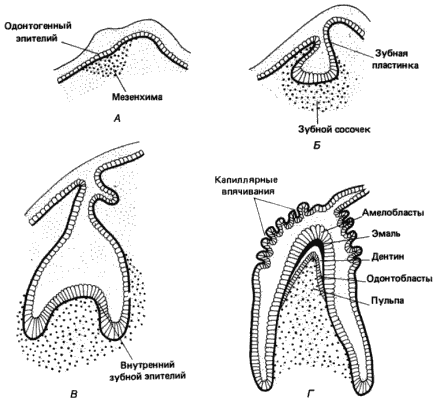
Рис. 5-7. Морфогенез зуба у млекопитающих. А. Слой эпителиальных клеток зубной пластинки, лежащий над оральной мезенхимой. Б. Эпителиальные клетки под влиянием индукционного сигнала со стороны мезенхимы наводняют последнюю. В. Зубная пластинка, индуцированная к образованию эмалевого органа, в свою очередь индуцирует дифференцировку мезенхимы в одонтобласты. Г. Формирование зуба в результате отложения дентина мезенхимными одонтобластами и эмали - эпителиальными одонтобластами.
У птиц в результате обычного взаимодействия между оральной мезенхимой и эпидермисом образуется клюв, а не зубы. Хаяши (Hayashi), используя гетероспецифичные комбинации оральных тканей курицы и утки, показал, что характер клюва определяется видовой принадлежностью мезенхимы. Так, при комбинации эпителия куриного зародыша с мезенхимой зародыша утки развивался клюв с характерными для утки зубчиками. Аналогичным образом Коллар и Байрд (Kollar, Baird) обнаружили, что оральный эпителий мыши поддается влиянию той мезенхимы, с которой он выращивается.
Наиболее показательный «эволюционный» эксперимент состоит в комбинации орального эпителия птицы с оральной мезенхимой млекопитающего. Коллар и Фишер (Fischer) провели этот эксперимент, выращивая кусочки глоточного эпителия курицы с молярной мезенхимой мыши. Обе ткани выращивали в передних камерах глаза половозрелых мышей, принадлежащих к особой генетической линии («голые» мыши), которые служат пусть несколько необычной, но подходящей культуральной средой. Против ожидания оральный эпителий курицы реагировал на оральную мезенхиму мыши, образуя структуры, похожие на эмалевые органы. В нескольких случаях, один из которых изображен на рис. 5-8, из этих кусочков развились настоящие зубы. Таким образом, в геноме по крайней мере одного вида птиц все еще сохраняется генетическая информация, дающая возможность оральному эпителию курицы успешно участвовать в последовательных взаимодействиях, необходимых для морфогенеза зубов и синтеза эмали. Поэтому утрату зубов у птиц можно рассматривать как результат изменения программы развития их мезенхимы, которое привело к выпадению начальных стадий этого процесса.
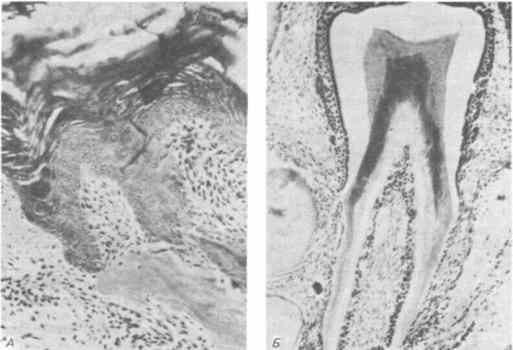
Рис. 5-8. Гибридный зуб, образовавшийся в результате совместного выращивания орального эпителия куриного зародыша и дентальной мезенхимы мышиного. А. Оральный эпителий куриного зародыша, образующий примитивный эмалевый орган, в котором находится мезенхима мышиного зародыша. Б. Хорошо развитый гибридный зуб (Kollar, Fisher, 1980).
Редукция малой берцовой кости у птиц также произошла в результате изменения программы развития, а не утраты генетической информации, необходимой для формирования этой кости. У археоптерикса имелась полностью развитая малая берцовая кость с суставными поверхностями на обоих концах; в отличие от этого у современных птиц (рис. 5-9) малая берцовая кость представляет собой всего лишь костный отросток, лежащий вдоль большой берцовой кости и приросший к ней. Исчерпывающее исследование взаимоотношений между костями ноги у куриного зародыша в процессе развития провел А. Ампэ (А. Hampe). Ампэ поставил ряд экспериментов, в которых различные участки развивающейся почки конечности были помечены введенными в них частичками угля. Местоположение этих меток в развитой конечности позволило Ампэ составить карту проспективных областей почки конечности (рис. 5-9). Главные такие области соответствуют скоплениям мезенхимных клеток, предназначенных для образования бедренной, большой и малой берцовых, плюсневых и предплюсневых костей. Ампэ проделал три типа экспериментов, убедившие его в том, что редуцированные размеры малой берцовой кости у современных птиц представляют собой результат захвата клеток, относящихся к проспективной области малой берцовой кости, проспективной областью большой берцовой кости.
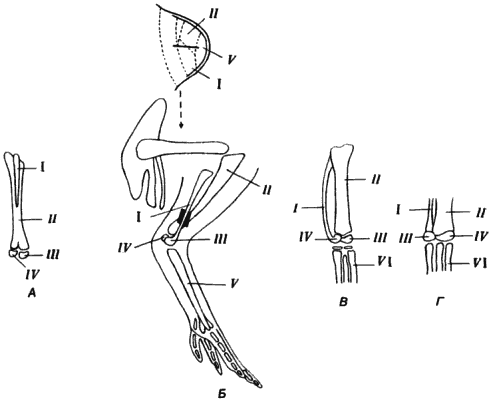
Рис. 5-9. Атавизм, проявляющийся в развитии ноги у курицы при экспериментальной модификации проспективной области костной ткани в почке конечности. А. Нормальная нога курицы. Б. Введение чешуйки слюды в почку конечности, между проспективными областями малой (I) и большой (II) берцовых костей и образующаяся в результате этого нога с увеличенной малой берцовой костью, несущей на дистальном конце суставную поверхность. В. Проявление атавизма в строении ноги курицы. Г. Нижняя конечность археоптерикса. III - fibulare; IV - tibiale; V - цевка; VI - плюсна (Hampe, 1959, 1960).
Первый из экспериментов, проведенных Ампэ, состоял в удалении или добавлении мезенхимных клеток к почке конечности. При удалении клеток из обеих берцовых костей конкуренция между ними усиливалась и малая берцовая кость не развивалась вовсе. Если же к почке конечности добавляли мезенхимные клетки, то большая берцовая кость не изменялась, а малая берцовая достигала одинаковых с нею размеров. Конкуренцию можно было подавить и другим способом: поворот проспективной области большой берцовой кости на 90° приводил к росту большой и малой берцовых костей в разных направлениях, так что конкуренция между ними становилась невозможной. В этом случае опять-таки малая берцовая достигала полной длины. Эти эксперименты ясно выявили конкуренцию между проспективными областями двух берцовых костей; однако наиболее яркие и интересные сведения об участвующих в этом эволюционных изменениях дал третий эксперимент.
Ампэ осторожно помещал между двумя проспективными участками почки конечности маленький кусочек слюды, который препятствовал проникновению презумптивных клеток малой берцовой кости в область большой берцовой. Полученный при этом неожиданный результат изображен на рис. 5-9: у сформировавшейся конечности малая берцовая не только достигала «полной длины», но и несла на своем дистальном конце суставную поверхность. Форма малой берцовой и ее взаимодействие с двумя маленькими предплюсневыми косточками, tibiale и fibulare, были очень близки к гомологичным структурам ноги археоптерикса. У взрослой курицы эти две косточки полностью срастаются с голенью. Разделение проспективных областей большой и малой берцовых костей, произведенное Ампэ, привело не только к образованию более длинной малой берцовой, но и к восстановлению древнего типа развития, который в течение долгого времени оставался подавленным. Гены, определявшие форму ноги у археоптерикса, все еще сохраняются у курицы, но в результате модификаций морфогенетических взаимодействий эти гены утратили возможность экспрессироваться, во всяком случае таким образом, как прежде.
Примеры регуляторных изменений, связанных с утратой зубов и редукцией малой берцовой кости у птиц, дают представление о тех способах, которыми регуляторные механизмы, оказывающие влияние на клеточные взаимодействия, могут вызывать эволюционные изменения морфологии. Однако оба этих примера имеют один недостаток: как в одном, так и в другом из них происходит редукция структуры. Если бы эволюция стремилась к своего рода морфологической нирване, это было бы превосходно, но морфологическая эволюция влечет за собой также модификации структур, а иногда и появление новых структур. К сожалению, случаи, когда эволюция приводит к утрате структур (как в двух уже рассмотренных примерах, а также в других, таких как утрата глаз у мексиканской пещерной рыбы, которую изучали Кан (Cahn) и Садоглу (Sadoglu), показавшие обусловленную генетически редукцию способности сетчатки индуцировать развитие у этих рыб хрусталика, легче поддаются экспериментальному анализу, чем случаи приобретения структур или их модификации. Эффектные превращения покровов, происходившие у рептилий, птиц и млекопитающих, повлекли за собой сложные изменения в системах регуляторных генов, и морфогенетические процессы, лежащие в основе развития чешуи, перьев и волос, требуют изучения. Хотя регуляция морфогенеза в этих системах все еще мало понятна, в общих чертах эволюционные изменения соответствующих регуляторных систем начинают выявляться благодаря работам П. Сенджела (Sengel) и его сотрудников, в особенности Дхуайи (Dhouailly).
Кожа состоит из двух слоев: верхнего - эпидермиса, происходящего из эктодермы зародыша, и лежащего под ним слоя дермы, происходящей из мезодермы. Развитие чешуи или других эпидермальных структур зависит от индукционных сигналов, исходящих от дермы.
Предполагается, что перья и волосы возникли в процессе эволюции из чешуи рептилий. Все эти структуры состоят из белков, принадлежащих к одному семейству кератинов. На тесную гомологию между чешуями и перьями указывает наблюдаемое иногда превращение кончиков чешуи, покрывающих ноги птиц, в перья.
Утиные перья сильно отличаются по своей морфологии от куриных. Сенджел и его сотрудники исследовали источник морфогенетической информации, необходимой для развития перьев, комбинируя зачатки дермы и эпидермиса, взятые от разных видов. Оказалось, что морфогенез пера детерминируется дермой. Общая архитектоника перьев, их размеры и число бородок соответствовали тому виду, от которого была взята дерма. Только форму клеток крючочков детерминировал эпидермис. Дерма детерминировала также характер распределения перьев. Как показали дальнейшие эксперименты с куриными зародышами, в которых дерму комбинировали с эпидермисом из области спины, дающим начало перьям, и с эпидермисом из области цевки, дающим начало характерным большим чешуям, такого рода детерминирующая роль дермы в морфогенезе широко распространена. Будет ли эпидермис детерминирован к образованию зачатков перьев или крупных чешуи, всегда зависело от типа морфогенеза, характерного для того участка зародыша, из которого брали дерму. Так, например, дорсальный эпидермис, обычно образующий перья, при комбинации с дермой из области цевки давал крупные чешуи.
И чешуи, и перья состоят из кератинов, однако они сильно различаются по набору содержащихся в них кератинов. При комбинировании спинных и тарзо-метатарзальных дермы и эпидермиса куриного зародыша у эпидермиса независимо от того, откуда он был взят, выбор экспрессируемого кератинового гена определяется дермой. Таким образом, расположение структур, образуемых эпидермисом, их морфологическая специфичность и экспрессия кератиновых генов детерминируются индукционными сигналами, поступающими от дермы. Остается, однако, открытым вопрос о том, как изменялись эти информационные сигналы в процессе эволюции высших позвоночных.
Гетероспецифичные комбинации возможны не только в пределах одного класса, но и между классами. Дхуайи провел замечательную серию экспериментов, соединяя в разных комбинациях дерму и эпидермис ящерицы, курицы и мыши. Как показали эти эксперименты, индукция слагается из двух этапов и эти этапы различаются в отношении степени их специфичности. Некоторые из полученных Дхуайи результатов представлены на рис. 5-10. Реакция эпидермиса ящерицы на куриную или мышиную дерму особенно поучительна. Эпидермис ящерицы реагирует на тарзо-метатарзальную дерму, образуя крупные чешуи, неотличимые по виду и расположению от чешуи на цевке птиц. Эпидермис ящерицы реагирует также на дорсальную дерму курицы, которая у кур детерминирует распределение и морфологию перьев. Однако эпидермис ящерицы неспособен интерпретировать специфические сигналы, вызывающие у кур образование перьев. Вместо перьев эпидермис ящерицы образует недоразвитые чешуи, расположенные в типичном для перьев гексагональном порядке. Точно так же эпидермис ящерицы в сочетании с мышиной дермой производит не волосы, а недоразвитые чешуи, расположенные типичным для шерстного покрова образом. Аналогичные картины наблюдаются при комбинации тканей курицы и мыши. Местоположение эпидермальных структур и ранние стадии их дифференцировки контролируются регуляторными веществами дермы, которые сохранились в процессе эволюции в таком виде, что эпидермальные клетки, принадлежащие представителям других классов, восприимчивы к их воздействиям. Для полной же дифференцировки зачатков чешуи, перьев или волос, очевидно, необходимы сигналы, специфичные для животных данного класса.
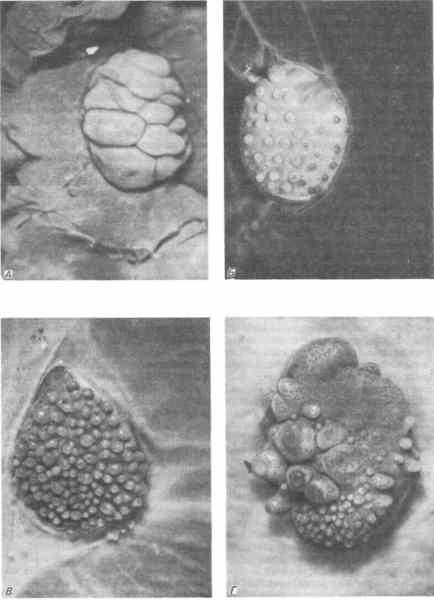
Рис. 5-10. Морфологические структуры, образующиеся при комбинациях эпидермиса ящерицы, который в норме дает ряды мелких чешуи, с дермой представителей других классов позвоночных в культуре органов. А. С куриной дермой из области цевки; формируются чешуи, расположенные так, как на цевке курицы. Б. Со спинной дермой курицы; формируются не перья, а недоразвитые чешуи, расположенные в типичном для перьев гексагональном порядке. В. Со спинной дермой мыши; формируются чешуи, расположение которых соответствует таковому первичных волосяных фолликулов. Г. С мышиной дермой из области верхней губы; формируются крупные чешуи, расположенные в соответствии с типичным расположением вибрисс и окруженные мелкими чешуями, расположенными подобно волосяным фолликулам шерстного покрова (Dhouailly, Sengel, 1973).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Ящерицы
Ящерицы Чаще всего попадаются на глаза и привлекают наше внимание ящерицы, которые доставляют одно из первых развлечений всякому пришельцу из холодных стран. Они буквально кишат повсюду. В городах видишь, как они гоняют по стенам и заборам, греются на солнце на сложенных
Подземный инкубатор глазчатой курицы
Подземный инкубатор глазчатой курицы Речь идет о так называемых большеногах, известных также под названием сорных кур. Ученым известно 12 видов этих птиц, область распространения которых ограничивается Австралией, Новой Гвинеей и прилежащими к ним островами
Крабы. Их враги. Мангусты. Лягушки и жабы. Крысы. Ящерицы
Крабы. Их враги. Мангусты. Лягушки и жабы. Крысы. Ящерицы По-видимому, всем людям случается рано или поздно столкнуться в жизни с чем-то таким, что им кажется не просто странным, а почти невероятным. Для тех, кто с этими явлениями уже сталкивался, они чаще всего становятся
Морские ящерицы ничего не боятся
Морские ящерицы ничего не боятся Жизнь рептилии протекает медленно. Рептилия как животное, меняющее температуру своего тела, не без помощи окружающей среды может поддерживать ее стабильной: если на улице холодно, ящерица тоже остывает, и ее обмен веществ работает в
Сухопутные крокодилы и ящерицы с «пастью дьявола»
Сухопутные крокодилы и ящерицы с «пастью дьявола» Пойманный дракон Никто не может поручиться, что когда-нибудь будут пойманы африканские «драконы», но хорошо известно, что «дракон» из Комодо — небольшого островка Индонезии — уже пойман.Но прежде послушайте, что
Часть 4 Любовь и курицы
Часть 4 Любовь и курицы Тихий ветер. Вечер сине-хмурый. Я смотрю широкими глазами. В Персии такие ж точно куры, Как у нас в соломенной Рязани. Тот же месяц, только чуть пошире, Чуть желтее и с другого края. Мы с тобою любим в этом мире Одинаково со всеми, дорогая. Сергей
Убийца в перьях
Убийца в перьях Быстро наступали сумерки, и я решил провести ночь не на дереве, а в пещере. Забаррикадировав вход принесенными из леса сучьями, я устроился на подстилке из мягкой душистой хвои и заснул. Рано утром я вышел и, спрятавшись за дюнами, наблюдал, как вереница