Филогения членистоногих, или как гомеология повторяет филогению
Филогения членистоногих, или как гомеология повторяет филогению
Филогенетические взаимоотношения крупных групп членистоногих представлены на рис. 8-10. Эта филогения построена на основании данных палеонтологических, эмбриологических, сравнительно-анатомических и различных других исследований.
Членистоногие возникли от некого предка, сходного с кольчецами, по всей вероятности, в конце докембрия, а первая значительная радиация этой группы началась в кембрии. Среди ископаемых остатков, найденных в нижнепалеозойских отложениях, обнаружены представители всех групп членистоногих, за исключением насекомых. Насекомые появляются в верхнем палеозое, т. е. примерно 350 млн. лет назад. Если в отношении происхождения членистоногих от кольчецов (или от какой-то сходной с ними группы) существует известное единодушие, то по вопросу о том, являются ли главные группы членистоногих моно- или полифилетическими, единого мнения нет. Мы не собираемся вступать в эту дискуссию, а сконцентрируем внимание на той части филогении, которая относится к группе членистоногих Uniramia и от которой в конечном счете произошли насекомые. Изменения в характере сегментации у этой группы членистоногих можно проанализировать в свете гомеозисных мутаций дрозофилы.
К признакам, позволяющим считать кольчецов предками членистоногих, относятся лежащая в основе их строения метаметрия, расположение нервной системы на вентральной стороне, а сердца - на дорсальной стороне тела; это признаки, которыми обладают все членистоногие. Несмотря на эти общие для обеих групп признаки взрослых особей, они сильно различаются по ранним стадиям развития. Для кольчецов, так же как для моллюсков, характерно спиральное дробление, тогда как у членистоногих дробление в общем центролецитальное, описанное выше для дрозофилы. Поэтому одним из крупнейших событий в возникновении членистоногих было резкое изменение раннего развития при сохранении взрослыми особями основного плана строения.
На рис. 8-11 показаны поздние стадии развития морской полихеты Polydora ciliata. После дробления и гаструляции у этого червя образуется свободноплавающая личинка - трохофора. Эта личинка разделена двумя венчиками ресничек на три обособленные области. Передняя, предротовая, часть отделена от средней, туловищной, части прототрохом. Задняя, пигидиальная, отделена от средней части телотрохом. В процессе развития личинки ее туловищная часть разделяется на три обособленных туловищных сегмента. На этой стадии у личинки можно различить в общей сложности шесть сегментов: предротовой, ротовой, три туловищных и хвостовой, или пигидий. Личинка продолжает расти путем добавления новых сегментов в зоне нарастания, находящейся между последним туловищным сегментом и пигидием. Этот рост кзади сопровождается постепенным усложнением структур, специфичных для каждого вида, на самых задних сегментах. После того как личинка достигнет полного развития, она в результате метаморфоза превращается во взрослого червя. Этот последний вопрос нас здесь не интересует; нам достаточно отметить сегментарную структуру животного и то, как эта сегментарная структура изменяется на «следующей» стадии филогенеза насекомых.
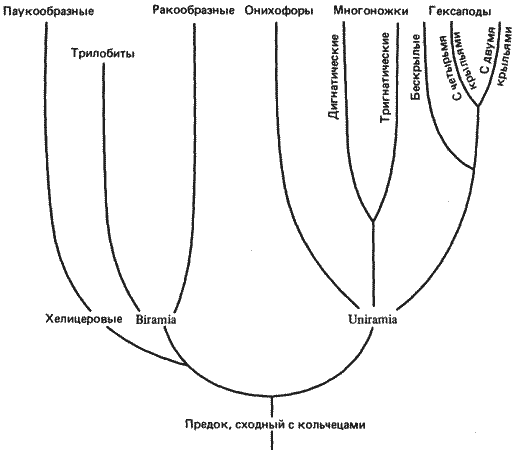
Рис. 8-10. Филогенетические связи между главными группами членистоногих с упором на линию, давшую начало современным насекомым (Hexapoda).
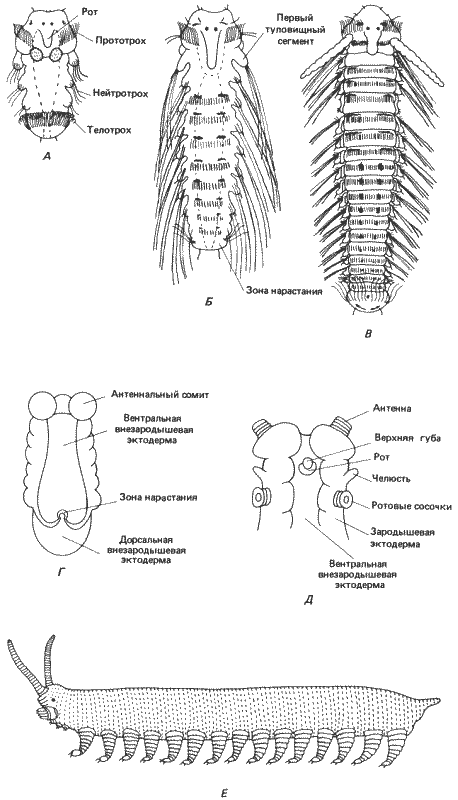
Рис. 8-11. А-В. Сегментация тела морской полихеты Polidora. А. Ранняя личиночная стадия. Б. Более поздняя личиночная стадия после добавления нескольких сегментов к зоне нарастания на заднем конце тела. В. Молодая взрослая особь. Г-Е. Зародыши и взрослая форма онихофор. Г. Вентральная сторона зародыша Peripatoides novaezealandiae на ранней стадии образования зародышевой полоски; как и у кольчецов, добавление новых сегментов происходит на заднем конце тела. Д. Вентральная сторона головы зародыша P. capensis, состоящей из трех сегментов - антеннального, челюстного и ротового - и головных сосочков. Е. Обобщенная взрослая онихофора (рисунок сделан с зафиксированных экземпляров); все туловищные сегменты одинаковы, и на каждом из них имеется по одной паре членистых ходильных ножек с коготком на конце (А-В - Wilson, 1928; Г - Sheldon, 1889; Д - Sedgwick, 1888).
Когда впервые были открыты онихофоры, их сочли «недостающим звеном» между кольчецами и высшими членистоногими. На первый взгляд это кажется правильным, и они в самом деле представляют собой промежуточную форму. Однако онихофоры не являются прямыми предками насекомых. Дробление у них центролецитальное, и у видов, имеющих крупные, богатые желтком яйца, бластодерма образуется на поверхности массы желтка путем целлюляризации. В процессе гаструляции на среднебрюшной поверхности бластодермы формируются две эктодермальные зародышевые полоски и происходит инвазия мезодермальных клеток в бластодерму. Крупные гаструляционные перемещения клеток возникают только в связи с образованием переднего зачатка средней кишки. Эти перемещения начинаются на презумптивном переднем конце животного и распространяются к хвостовому его концу. Вслед за началом этих перемещений начинается процесс сегментации. Ранняя стадия сегментации Peripatoides novaezealandiae показана на рис. 8-11, Г. На переднем конце животного имеется хорошо заметный антеннальный сегмент, за которым на этой стадии следует ряд сходных сегментов, дающих сомиты. Как и у личиночных форм полихет, новые сегменты образуются субтерминально, в задней зоне нарастания. Сегментация завершается до рождения на свет (большинство видов онихофор живородящие или яйцеживородящие). На рис. 8-11, Д изображена более поздняя стадия эмбриогенеза Peripatopsis capensis, соответствующая началу развития головы. Голова состоит из переднего антеннального сегмента, единственного челюстного сегмента, на котором находится рот, и сегмента, несущего ротовые сосочки, где открываются протоки слизистых желез. Позади головы расположен ряд одинаковых туловищных сегментов, от каждого из которых у взрослой особи отходит членистая нога, снабженная коготком.
Антеннальный, челюстной и сосочковый сегменты - цефализированные туловищные сегменты, которые мобилизованы, в эволюционном и онтогенетическом смысле, функционировать в качестве ротовых частей. Надежных данных о наличии у онихофор процефалического сегмента, подобного имеющемуся у кольчецов и высших Uniramia, нет (табл. 8-1). Отчасти поэтому современные онихофоры считаются реликтами, а не подлинным промежуточным звеном между кольчецами и следующей ступенью в филогенезе насекомых.
Таблица 8-1. Соответствие между сегментами кольчецов и членистоногих, принадлежащих к группе Uniramia
Номер сегмента Кольчецы Онихофоры Многоножки Гексаподы Дигнатические Тригнатические Бескрылые Крылатые 1 Предротовой Антеннальный Процефалический Процефалический Процефалический Процефалический 2 Ротовой Ротовой Антеннальный Антеннальный Антеннальный Антеннальный 3 Метатрохальный Челюстной Премандибулярный Премандибулярный Премандибулярный — 4 Туловищный Сосочковый Мандибулярный Мандибулярный Мандибулярный Мандибулярный 5 ' Туловищный Максиллярный Максиллярный Максиллярный Максиллярный 6 ' ' Шейный Нижнегубной Нижнегубной Нижнегубной 7 ' ' Туловищный Туловищный Грудной 1 Грудной 1 8 ' ' ' ' Грудной 2 Грудной 2 9 ' ' ' ' Грудной 3 Грудной 3 10 ' ' ' ' Брюшной 1 Брюшной 1 11 ' ' ' ' ' ' 12 ' ' ' ' ' ' 13 ' ' ' ' ' ' 14 ' ' ' ' ' ' 15 ' ' ' ' ' ' . ' ' ' ' ' ' . ' ' ' ' ' ' n ' ' ' ' ' 'У многоножек - очередной ступени филогенетической лестницы, по которой мы следуем,- имеются разного рода второстепенные изменения общего плана развития, наблюдаемого у онихофор. Дробление у них продолжает оставаться центролецитальным, хотя у некоторых групп гаструляция несколько изменена. Многоножки делятся на две основные группы - дигнатические и тригнатические - различающиеся по сегментации головы (табл. 8-1, рис. 8-10). Возможно, что дигнатические многоножки появились в процессе эволюции позднее, несмотря на менее сложное строение их головы. После гаструляции зародышевая полоска пересекается рядом борозд, в результате чего образуется группа сегментов: у тригнатических многоножек - это головные, антеннальные, премандибулярные, мандибулярные, максиллярные. нижнегубные и от 3 до 6 туловищных сегментов. Эта стадия показана на примере Hanseniella (на рис. 8-12, Д и Е). Как и у онихофор, полный комплект сегментов взрослой особи создается в процессе эмбриогенеза (в зоне нарастания между предпоследним задним сегментом и анальной областью). Дальнейшее развитие приводит к совершенствованию строения ходильных ног, имеющихся на каждом из туловищных сегментов, и к формированию ротовых частей из мандибулярного, максиллярного и нижнегубного сегментов (рис. 8-12, Ж и 3). Премандибулярный сегмент несет лишь временный придаток и не играет заметной роли в формировании головы взрослой особи. Лежащие впереди антеннальный и процефалический сегменты несут антенны и клипеолабрум соответственно. Таким образом, у многоножек в образовании головы участвуют не три, как у онихофор, а шесть сегментов. Но, так же как и у онихофор, эти специализированные сегменты возникли в результате заимствования элементов из послеротовой туловищной области раннего зародыша. У дигнатических многоножек наблюдается своеобразное отличие в типе сегментации. Сегмент, соответствующий нижнегубному сегменту тригнатических многоножек, не участвует в образовании ротовых частей взрослой особи. Вместо этого он превращен в специализированный шейный сегмент (collum), образуя подобие шейки между головой и первым туловищным сегментом (рис. 8-12, А и Б). В остальном характер сегментации у обеих групп многоножек сходен. В частном примере Pauropus silvaticus, представленном на рис. 8-12, А-Г, есть еще одно резкое и существенное различие. Для класса Pauropoda, к которому относится эта многоножка, характерен анаморфоз, а не эпиморфоз, как у описанного выше представителя Chilopoda. При развитии с анаморфозом животное вылупляется с неполным набором туловищных сегментов. Как видно на рис. 8-12, В, у только что вылупившейся особи имеются всего три туловищных сегмента. Остальные сегменты, имеющиеся у взрослой особи, приобретаются в результате субтерминального их добавления в зоне нарастания на заднем конце животного. Это нельзя считать признаком дигнатизма как такового, потому что у Symphyla - группы тригнатических многоножек - также наблюдается анаморфоз. Существование личиночной формы, имеющей всего три туловищных сегмента, было сочтено частичным доказательством того, что насекомые возникли от какого-то сходного с многоножками предка в результате педогенеза. В такой возможности убеждает изучение примитивных бескрылых насекомых (Apterygota).
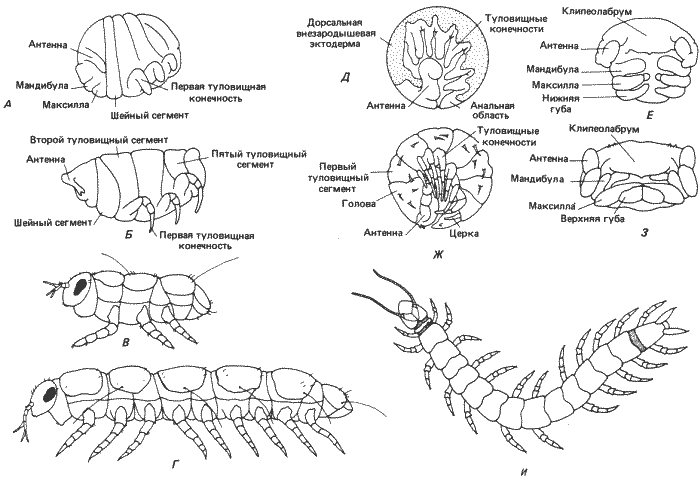
Рис. 8-12. Зародышевая, личиночная и взрослая стадия (А-Г) дигнатической многоножки Pauropus, зародышевые стадии тригнатической многоножки Hanseniella (Д-3) и взрослая тригнатическая многоножка Scutigerella (И). А. Десятидневный зародыш с намечающейся сегментацией тела. Б. Стадия ранней куколки. В. Личинка первого возраста вскоре после вылупления. Г. Взрослая особь после добавления туловищных сегментов в зоне нарастания на заднем конце тела. Д. Вся зародышевая полоска 8-дневного зародыша (вид сбоку). Е. Ротовые части того же зародыша, что на Д (вид снизу). Ж. Зародыш непосредственно перед вылуплением (вид сбоку). 3. Голова и ротовые части того же зародыша, что на Ж (вид снизу). И. Взрослая особь (вид сверху). (А, Б, Д-3 - Anderson, 1973; В, Г, И - по Snodgrass, 1952; с изменениями).
Среди бескрылых насекомых есть группы, раннее развитие которых сходно с развитием либо многоножек, либо крылатых насекомых. Андерсон (Anderson) считает эти вариации функциональными адаптациями каждой отдельной группы, а не существенными различиями, опровергающими наличие связи между многоножками и насекомыми. Наиболее ярко выраженное сходство между многоножками, бескрылыми и крылатыми насекомыми и между самими бескрылыми касается способа сегментации зародышевой полоски. У бескрылых, так же как и у тригнатических многоножек, в начале процесса сегментации имеется шесть головных сегментов, за которыми следуют три туловищных сегмента. В процессе дальнейшего развития в субтерминальной зоне нарастания кпереди от терминального хвостового сегмента образуются остальные туловищные сегменты (рис. 8-13, Г и Д). Гнатоцефалические сегменты пополняются за счет туловищных элементов зародыша и их придатков, модифицированных в ротовые части. Главное отличие бескрылых от многоножек состоит в том, что их туловище разделено на грудь, состоящую из трех сегментов, и брюшко из восьми сегментов. Ходильные ноги имеются только на груди, а придатки брюшка сильно редуцированы (рис. 8-13, Е). Сравнение рис. 8-12, В и Г и 8-13, Е выявляет поразительное сходство в строении тела анаморфных многоножек и примитивных бескрылых насекомых; главное различие между ними - подавление у бескрылых насекомых развития конечностей на сегментах, расположенных позади третьего туловищного сегмента.
Последняя ступень в филогенезе - крылатые насекомые (Pterygota). Мы уже рассматривали некоторые аспекты раннего развития этой группы на примере Drosophila melanogaster (см. рис. 8-6). События раннего развития, описанные для дрозофилы, наблюдаются у всех других крылатых насекомых, и нет нужды повторять их. В качестве примера более примитивной формы мы сочли нужным, однако, рассмотреть сегментацию зародышевой полоски у жука Bruchidius (рис. 8-13). Так же как и у дрозофилы, у этого жука, по-видимому, нет задней зоны нарастания и сегментация происходит единовременно. У него имеются три гнатоцефалических и три грудных сегмента, образующие соответственно ротовые части и грудь. В отличие от бескрылых насекомых три грудных сегмента у него дифференцированы, причем от второго и третьего сегментов отходят крылья.
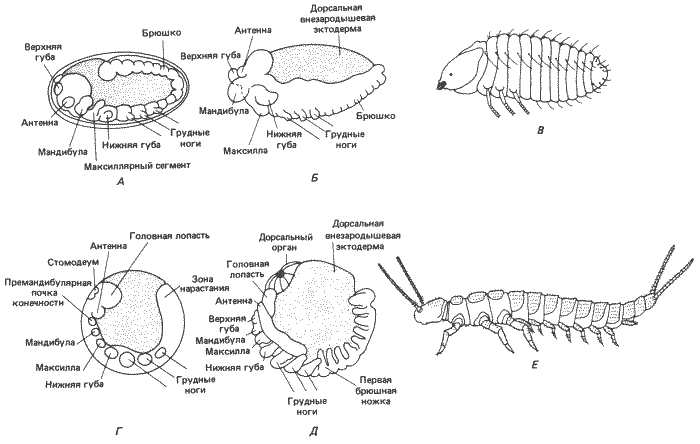
Рис. 8-13. Вид сбоку сегментированных зародышей и вылупившихся особей крылатого насекомого Bruchidius (А-В) и бескрылого Campodea (Г-?). А. Полностью сегментированная зародышевая полоска. Б. Укорочение зародышевой полоски. В. Стадия вылупления. Г. Сегментирующаяся зародышевая полоска; сегментация происходит на заднем конце полоски. Д. Полностью сформировавшаяся и сегментированная зародышевая полоска. Е. Взрослая особь. При сравнении рис. В и ? с многоножками, изображенными на рис. 8-12, В и Г, бросается в глаза сильное сходство между этими формами, которое отчасти и послужило основанием для гипотезы о происхождении примитивных насекомых в результате педогенеза от предка, сходного с многоножками (А-В - Anderson, 1973; E - Snodgrass, 1952).
У двукрылых произошли дальнейшие изменения: вторая пара крыльев, отходящая от заднегруди, редуцировалась, превратившись в жужжальца, а спинка переднегруди сильно уменьшена и у дрозофилы сведена к узкой полоске кутикулы (см. рис. 8-7). Брюшные конечности, которые у бескрылых редуцированы, у всех крылатых насекомых совершенно отсутствуют. Кроме того, премандибулярный головной сегмент у них плохо различим или отсутствует вовсе. Эта редукция головных сегментов заходит еще дальше у дрозофилы, у которой мандибулярный сегмент участвует в формировании ротовых частей личинки, но не имаго. На рис. 8-14 суммированы изменения сегментов, наблюдаемые у Uniramia. Следуя по филогенетической лестнице от кольчецов до крылатых насекомых, можно видеть постепенное возникновение сегментированной структуры и последующую утрату некоторых, но не всех ее элементов. Сравнение рис. 8-14 и 8-9 позволяет провести довольно интересную параллель. Выпадение функции генов bx+ и pbx+ превращает двукрылую дрозофилу в более примитивную четырехкрылую форму. Функция гена Antp+ необходима для развития средне-и заднегрудных сегментов. В отсутствие этого гена грудь зародыша состоит из трех переднегрудных сегментов, которые не несут крыльев, т.е. частично подражают бескрылым формам. В отсутствие всего комплекса ВХ-С задний конец зародыша состоит из ряда одинаковых грудных или туловищных сегментов. Такой набор сегментов напоминает тригнатических многоножек. Наконец, в отсутствие комплексов ANT-C и ВХ-С развиваются зародыши с тремя головными сегментами и рядом одинаковых туловищных сегментов, что напоминает строение онихофор. Таким образом, постепенно удаляя относительно небольшое количество генетического материала, нам удалось пройти довольно большой отрезок эволюционного пути.
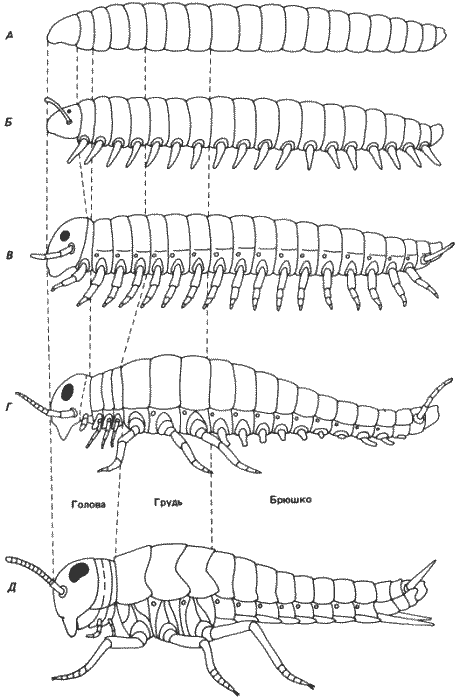
Рис. 8-14. Схема предполагаемой эволюции характера сегментации насекомых в процессе филогенеза. А. Кольчецы. Б. Онихофоры. В. Многоножки. Г. Бескрылые насекомые. Д. Крылатые насекомые. Сопоставляя эту схему с рис. 8-9, можно видеть, что постепенное выпадение гомеозисных локусов приводит к прогрессирующему упрощению сегментации у дрозофилы, которое до некоторой степени имитирует филогению насекомых. (Snodgrass, 1935).
Мы не хотели бы, чтобы читатель принял эти аналогии слишком буквально, и считаем необходимым подчеркнуть, что описанные здесь атавизмы нельзя считать подлинными. Делеция генов привела лишь к изменению индивидуальности некоторых сегментов, но при этом они несомненно остаются сегментами дрозофилид. Это показывает, что содержащиеся в этих локусах гены участвуют в спецификации развивающейся структуры - в регуляции онтогенеза как такового. Они играют роль переключателей, определяя судьбу клеток путем регуляции действия генов, экспрессирующихся в каждом отдельном сегменте. Находящиеся под их контролем структурные гены, по-видимому, эволюционировали совместно с этими контролирующими гомеозисными локусами, но в то же время в известном смысле обособленно от них.
На основе описанной здесь филогении насекомых и тонкой структуры двух комплексов генов мы представляем себе эволюцию гомеозисных форм как ступенчатую дупликацию с последующей дивергенцией функций. Самый подходящий отрезок филогенеза, где легче всего наблюдать этот процесс, - переход от многоножек к бескрылым и далее к крылатым насекомым. Гены iab определяют дифференцировку груди и брюшка. В отсутствие этих генов брюшные сегменты становятся грудными. Поэтому у генов iab возникла функция, которая могла подавлять развитие конечностей у задних сегментов. Льюис (Lewis) показал, что у дрозофилы имеется несколько таких локусов, возможно, по одному на каждый из восьми брюшных сегментов.
Однако у бескрылых насекомых эти сегменты добавляются один за другим в процессе эмбриогенеза; и возможно, что им для этого нужен только один ген, который многократно активируется по мере добавления новых брюшных сегментов. С возникновением типичного для крылатых одновременного формирования всех сегментов могла произойти серийная дупликация гена iab как необходимый элемент такого изменения развития.
Можно считать, что члены комплекса ANT-C возникли в какой-то ранний момент эволюции насекомых. Переход от предка, сходного с кольчецами, к онихофорам, а затем к многоножкам был связан с вовлечением послеротовых туловищных сегментов и превращением их в гнатоцефалические сегменты, дающие начало ротовым частям. Этот процесс осуществляется, по крайней мере частично, локусом proboscipedia. Кроме того, гены Scr+ и Antp+ (см. рис. 8-8, А) функционируют как переключатели, необходимые для детерминации характерных черт (индивидуальности) первого и второго грудных сегментов. Можно представить себе, что функции этих генов возникли, дуплицировались и дивергировали во многом таким же образом, как функции локусов ВХ-С. Возможно даже, что эти два комплекса связаны между собой через дупликацию: элементы комплекса ANT-C дали начало путем дупликации какому-то предшественнику комплекса ВХ-С.
До сих пор мы рассматривали типы трансформаций, вызываемых гомеозисными генами, а также место и сроки функционирования этих генов, не касаясь того, как они осуществляют переключение. Этот последний вопрос фактически состоит из двух частей: как регулируются сами гомеозисные гены и как они в свою очередь регулируют характер генной экспрессии в пределах каждого сегмента? Прежде чем попытаться ответить на эти два вопроса, необходимо обсудить существующие представления о способе становления пространственной структуры.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
§ 22. Нервная система членистоногих
§ 22. Нервная система членистоногих Организация нервной системы членистоногих и сходных с ними групп может существенно варьировать, но в пределах общего плана строения. Рисунок нервной системы дневной бабочки (Lepidoptera) довольно точно отражает типичное расположение
Происхождение членистоногих — «артроподизация» (вендский и кембрийский периоды)
Происхождение членистоногих — «артроподизация» (вендский и кембрийский периоды) В «домолекулярную эру» в распоряжении ученых было три научных дисциплины, при помощи которых можно было реконструировать эволюционную историю организмов: 1. сравнительная анатомия, 2.
Систематика и филогения
Систематика и филогения Если первые системы организмов были искусственными, т. е. основанными на произвольно выбранных признаках, то современная систематика строится на принципах филогении.Филогения – это историческое развитие организмов. Графически филогения