Нормальная сегментация у дрозофилы
Нормальная сегментация у дрозофилы
После оплодотворения и слияния гамет у зародыша дрозофилы наблюдается ряд синцитиальных делений дробления, т.е. деления ядер, не сопровождающиеся образованием клеток зародыша. Первые девять таких делений происходят по всей цитоплазме яйца, и после девятого деления большая часть ядер мигрирует в кортикальную цитоплазму. За этим следуют еще четыре деления ядер, и после завершения 13-го деления происходит целлюляризация, приводящая к образованию бластодермы, состоящей примерно из 6000 клеток. Эта стадия изображена на рис. 8-6, на котором соматические клетки бластодермы можно отличить по их морфологии от сферических полярных клеток (презумптивные клетки зародышевого пути), расположенных на заднем конце зародыша. Именно на этой стадии развития детерминируется судьба клеток, т.е. определяется их назначение как в личиночных, так и в имагинальных структурах. Кроме того, именно в этот момент геном зиготы впервые начинает проявлять заметную транскрипционную активность. Во время гаструляции на переднем конце зародыша образуется латеральная складка, называемая головной бороздой. Эта борозда отделяет большую часть головы от туловища. Одновременно происходит перенос полярных клеток с заднего конца зародыша на его дорсальную поверхность путем удлинения зародышевой полоски. В результате образования трех отдельных выпячиваний - вентральной борозды, заднего и переднего зачатков средней кишки - мезодерма и передняя и задняя энтодерма перемещаются внутрь зародыша. Вслед за этим, примерно на 8-й час развития, поверхность зародышевой полоски покрывается рядом латеральных складок, которые физически разделяют зародыш на сегменты. Сегменты зародыша можно видеть на рис. 8-6; они соответствуют метамерным сегментам личинки и имаго. Таким образом, после развития в течение 10 ч зародыш состоит из трех зародышевых листков и разделен на голову, грудь и брюшко. Голова состоит из клипеолабрального, процефалического, мандибулярного, максиллярного и нижнегубного сегментов. Три последних сегмента объединяют под названием гнатоцефалических. Грудь состоит из трех сегментов: переднегруди, среднегруди и заднегруди. Остальные девять сегментов делятся на восемь брюшных и один концевой хвостовой сегмент.
Эмбриогенез завершается созданием этого основного метамерного типа строения, образованием кутикулярных покровов и внутренних органов личинки. Личинка вылупляется через 24 ч после оплодотворения яйца. Передний вентральный край каждого из трех грудных и восьми брюшных сегментов окаймлен рядами зубчиков, форма которых позволяет отличать брюшные сегменты личинки от грудных. Сегменты головы инвертированы таким образом, что голова оказывается внутри личинки, где гнатоцефалические сегменты дают начало ротовым частям личинки. Зачатки имагинальных структур, или имагинальные диски, которые детерминируются и на стадии клеточной бластодермы отделяются от клеток, дающих начало тканям личинки, растут на протяжении трех личиночных стадий, а затем достигают своего взрослого дифференцированного состояния во время метаморфоза на стадии куколки.
Взрослая особь подобно зародышу и личинке, имеет метамерное строение. Различные имагинальные диски, очевидно, происходят из клеток, распределенных по разным метамерам зародыша. На рис. 8-7 схематически показано происхождение разных частей имаго из метамеров зародыша.
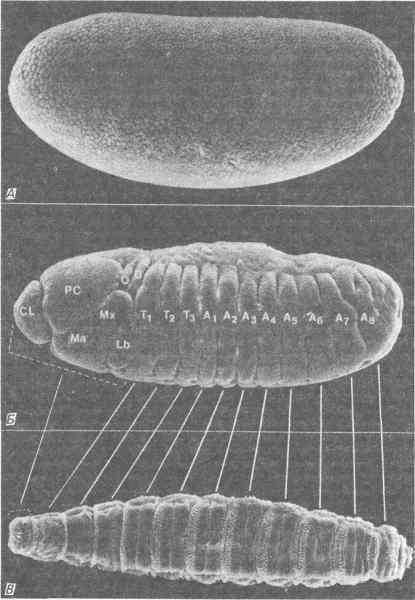
Рис. 8-6. Бластодерма (A), стадия сегментированной зародышевой полости (Б) и личинка первого возраста (В) Drosophila melanogaster. Микрофотографии, полученные с помощью сканирующего электронного микроскопа. Линиями соединены соответствующие друг другу сегменты 8-часового зародыша и сформировавшейся личинки. Головы зародыша и личинки показаны прерывистой скобой. Обозначения сегментов: CL - клипеолабральный; РС - процефалический; Ма - мандибулярный, Мх - максиллярный; Lb - нижнегубной; О - глазной; T1-Т3 - первый, второй и третий грудные; A1-A8 - от первого до восьмого брюшные; D - дорсальный валик. (Фотография любезно предоставлена д-ром F. R. Turner.)
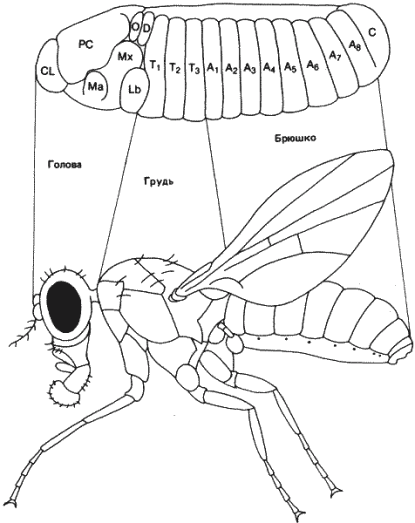
Рис. 8-7. Схема, показывающая соответствие между сегментами 8-часового зародыша и имаго Drosophila melanogaster (обозначения те же, что на рис. 8-6).
Голова взрослой особи несет глаза, антенны, клипеолабрум и максиллярные и нижнегубные щупики. Три последних элемента ротовых частей имаго, вероятно, происходят из соответствующих сегментов зародыша. У высших двукрылых, к которым относится и дрозофила, на имагинальной стадии мандибулярный элемент отсутствует. Глаза и антенны, по всей вероятности, происходят из процефалической лопасти зародыша. Вся голова имаго развивается из трех дисков, причем большая часть головной капсулы формируется из глазо-антеннального диска, который дает также начало максиллярным щупикам, т. е. этот диск происходит из нескольких сегментов. Два других диска происходят каждый из одного сегмента. Все три грудных сегмента несут по паре ходильных ног, причем каждая пара происходит из одного диска. Диски ног имеются на каждом из трех грудных сегментов зародыша. Дорсальная сторона груди имаго образована в основном мезотораксом и происходит из крылового диска. Дорсальные части как про-, так и метаторакса сильно редуцированы. На метатораксе расположены уже упоминавшиеся жужжальца. Наконец, восемь брюшных сегментов и концевые половые структуры имаго происходят из соответствующих сегментов личинки. Происходят ли половые железы из хвостового сегмента, не известно.
Все границы между сегментами появляются одновременно во время гаструляции. Установлено, однако, что процесс сегментации не абсолютно мозаичен по своему характеру. Эксперименты Шубигера и Вуда (Schubiger, Wood), а также Херта и Сандера (Herth, Sander) с наложением лигатуры на зародышей, находящихся на разных стадиях дробления, показали, что становление структуры сегментов с течением процесса дробления постепенно усиливается. Полный набор сегментов образуется лишь по достижении стадии бластодермы, когда настает время детерминации судьбы разных клеток. Очевидно, для нормальной сегментации зародыша необходимо взаимодействие между его различными участками. Дальнейшие сведения о природе факторов, определяющих характер сегментации, дали эксперименты, в которых клетки и их потомков метили, вызывая в них путем рентгеновского облучения соматические рекомбинации. Используя клеточные маркеры, экспрессирующиеся независимо друг от друга и затрагивающие пигментацию и морфологию кутикулярных элементов взрослой особи, Гарсиа-Беллидо (Garcia-Bellido) и его сотрудники создавали на стадии клеточной бластодермы клоны рекомбинантных клеток в зачатках, дающих грудные структуры взрослых особей. Проследив за взаимоотношениями между клеточными линиями, они сумели показать, что все потомки первоначально помеченной клетки оказываются только в одном сегменте. Более того, клетки, происходящие от одной гомозиготной дочерней клетки, обычно остаются друг подле друга, образуя пятно меченых клеток. Как показали дальнейшие эксперименты с использованием ? (Minute)-мутаций, границы между такими участками отражают ограниченность клеточных потенций, и сегменты в самом деле делятся на два таких ограниченных участка. Клетки, гетерозиготные по доминантной мутации ? (?/?+), растут медленнее, чем гомозиготные клетки дикого типа (?+/?+), а гомозиготные клетки по мутации М (М/М) гибнут. Если клоны клеток М+/М+ индуцировать при помощи соматической рекомбинации на фоне клеток ?/?+, то эти нормальные клетки обгоняют в росте своих гетерозиготных соседей. В больших клонах клеток М+/М+ проявляются те же ограничения потенций, которые наблюдались в предыдущих экспериментах. Итак, три сегмента, из которых состоит грудь имаго, разделены на переднюю и заднюю области («компартменты»). Все эти эксперименты с клонированием проводились на кутикулярных элементах взрослых мух. Перенесение полученных при этом результатов на зародышей и личинок до некоторой степени произвольно. Однако проведенный Корнбергом (Kornberg) анализ мутантных аллелей локуса еп (engrailed) подтверждает такую возможность. Leaky-мутации локуса еп вызывают превращение элементов задних компартментов грудных сегментов в структуры передних компартментов. Это особенно ясно выражено в случае имагинальной пластинки крыла, которая у мутанта состоит из двух передних половинок, зеркально-симметричных одна другой. Другие аллели вызывают гибель зародыша на поздних стадиях развития. У этих мертвых особей обнаружены сходные зеркально-симметричные дупликации всех грудных и брюшных сегментов. Поэтому создается впечатление, что компартментализация происходит уже у зародыша и что это распределение клеточных потенций внутри сегмента имеет место как у личинки, так и у взрослой особи.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
14. Нормальная микрофлора человека
14. Нормальная микрофлора человека Нормальная микрофлора человека – это совокупность множества микробиоценозов, характеризующихся определенными взаимосвязями и местом обитания.Виды нормальной микрофлоры:1) резидентная – постоянная, характерная для данного
ЛЕКЦИЯ № 7. Нормальная микрофлора организма человека
ЛЕКЦИЯ № 7. Нормальная микрофлора организма человека 1. Нормальная микрофлора человека Нормальная микрофлора человека – это совокупность множества микробиоценозов, характеризующихся определенными взаимосвязями и местом обитания.В организме человека в соответствии
1. Нормальная микрофлора человека
1. Нормальная микрофлора человека Нормальная микрофлора человека – это совокупность множества микробиоценозов, характеризующихся определенными взаимосвязями и местом обитания.В организме человека в соответствии с условиями обитания формируются биотопы с
ГЛАВА 4 НОРМАЛЬНАЯ ЩЕННОСТЬ
ГЛАВА 4 НОРМАЛЬНАЯ ЩЕННОСТЬ Беременность (щенность), равно как и роды (щенение), а также весь период выкармливания щенков под сукой — весьма ответственное время в жизни собаки и ее владельца. Это отдельная, большая тема для разговора, имеющая много аспектов, обсуждение
Нормальная беременность
Нормальная беременность На ранних стадиях невозможно определить, будет ли сука иметь щенков или нет, ни по внешнему виду, ни при прощупывании.Возможно, что первые признаки проявляются не в изменении физического состояния, а в поведении суки. Очень часто после вязки сука
НАСКОЛЬКО ПАРАНОРМАЛЬНА «НОРМАЛЬНАЯ» НАУКА?
НАСКОЛЬКО ПАРАНОРМАЛЬНА «НОРМАЛЬНАЯ» НАУКА? Для общепринятого табу на парапсихологию, делающего ее изгоем среди наук, есть веская причина. Существование некоторых парапсихических явлений могло бы серьезно подорвать веру в иллюзию объективности. Оно могло бы
Нормальная беременность
Нормальная беременность На ранних стадиях невозможно определить, будет ли сука иметь щенков или нет, ни по внешнему виду, ни при прощупывании.Возможно, что первые признаки проявляются не в изменении физического состояния, а в поведении суки. Очень часто после вязки сука
Бесплодные мыши и дрозофилы с красными глазами
Бесплодные мыши и дрозофилы с красными глазами Около десяти лет назад эпигенетику Ренато Паро удался захватывающий эксперимент. Он вывел мух с красными, а не белыми глазами, и все из-за воздействия теплового удара в эмбриональной фазе развития. В общем-то, ничего
Рассказ Дрозофилы
Рассказ Дрозофилы В 1894 году Уильям Бэтсон, один из отцов генетики, опубликовал книгу “Материалы по изучению прерывистой изменчивости в происхождении видов”. Он собрал впечатляющий, немного жутковатый список генетических отклонений и рассмотрел их с точки зрения