Выбор фенотипа и морфогенетический потенциал генома
Выбор фенотипа и морфогенетический потенциал генома
В регуляции развития со стороны генома есть еще один важный аспект. Фенотипы не строго детерминированы единой генетической программой. На морфологии сказываются также негенетические влияния среды, хотя, как это подробно рассматривает Уоддингтон, возможный диапазон фенотипических реакций детерминирован характером генетической программы. И в самом деле, выбор из альтернативных фенотипов представляет собой почти постоянный элемент любого онтогенеза. Фенотипическая пластичность - очень обширная и сложная тема; поэтому мы ограничимся здесь проблемой выбора между альтернативными фенотипами, что составляет часть программы развития вида и служит основой для эволюционных изменений, в чем можно убедиться на некоторых классических примерах гетерохронии. К числу явлений, в которых участвует запрограммированный выбор между фенотипами, относятся половой диморфизм, различие в морфологии личинок и взрослых особей и альтернативные типы морфологии взрослых особей, такие как определяемая условиями среды неотения у амфибий или различные касты у общественных насекомых, обусловленные гормонально. Во всех этих случаях выбор одной из альтернативных программ морфологического развития из всего репертуара возможностей данного генома производят переключатели.
Потенции генома не ограничиваются созданием одной неизменной морфологии; в этом можно убедиться на прекрасном и ярком примере разнообразия каст у термита Amitermes atlanticus. Как у большинства общественных насекомых, семья этих термитов основывается царицей, или маткой, после ее брачного полета. Рожденные царицей потомки образуют популяцию термитов данной колонии. В своей книге «Обитающие во тьме» Скайфе (Skaife) описал различные касты, которые можно обнаружить в зрелой колонии этих организмов: рабочие, солдаты, царицы (или матки) и самки-заменительницы. Представители всех этих каст изображены на рис. 9-9. Как можно видеть на этом рисунке (внизу), особи, относящиеся к разным кастам, сильно различаются по своей морфологии. Глаза и крылья имеются только у половых особей, тогда как рабочие и солдаты слепые и бескрылые. Солдаты отличаются от рабочих и половых особей крупной головой и мощными челюстями. Когда представители всех этих каст вылупляются из яиц, никаких различий между ними заметить нельзя. Следует также помнить, что все члены данной колонии - родные братья и сестры. Примерно в середине периода роста между развивающимися термитами начинают появляться морфологические различия, зависящие от их будущей касты. Вряд ли можно предположить, что эти различия обусловлены генетически, потому что молодые семьи состоят из одних только рабочих, а по мере созревания семьи в ней появляются сначала солдаты, а затем разные категории половых особей. Кроме того, в зрелой семье наблюдаются сезонные колебания числа и категорий половых особей. Таким образом, создается впечатление, что в группе организмов, приходящихся друг другу братьями и сестрами, т.е. имеющих один и тот же генотип, могут возникнуть довольно разнообразные фенотипы, причем все эти фенотипы принадлежат функционально интегрированным организмам.
Несколько лучше известны - главным образом благодаря работам Люшера (Luscher) - механизмы кастовой дифференциации у более примитивных термитов, обитающих под землей. Кастовая дифференциация у этих видов регулируется феромоном, подавляющим образование самок-заменительниц, и главными гормонами насекомых, действующими на развитие. По-видимому, превращение недифференцированных нимф в половых особей происходит под действием экдизона, тогда как высокие уровни ювенильного гормона вызывают превращение нимф в солдат. Таким образом, при эндокринной регуляции кастовой дифференциации, возможно, имеют место конкурентные взаимодействия экдизона и ювенильного гормона.
Роль гормонов в управлении морфогенетическими переключателями при развитии насекомых иллюстрируется далее работами Уилера и Ниджхаута (Wheeler, Nijhout), изучавших развитие солдат у муравья Pheidole bicarinata. Так же как у термитов, в семьях этих муравьев имеются рабочие, солдаты и половые особи. Оказалось, что солдат можно получить из недифференцированных личинок женского пола, если в определенный период последней личиночной стадии обработать их аналогом ювенильного гормона насекомых. Такая обработка приводит к увеличению продолжительности периода роста и изменению размеров личинок ко времени метаморфоза. Эти более крупные особи с более длительным периодом роста и есть солдаты. Как показали Дж. Гексли (J. Huxley) и Вилсон (Wilson), у муравьев аллометрические кривые рабочих особей и солдат совпадают (см. рис. 2-10). Огромные размеры челюстей и головы у солдат - следствие их более крупных общих размеров. У некоторых видов муравьев ситуация более сложная, потому что наклон аллометрических кривых изменяется с увеличением размеров. Известны двух- и трехфазные аллометрические зависимости. Изменение уровней гормонов позволяет установить пороги, с помощью которых муравьиная семья может «выбирать» морфологию, которую приобретает данная личинка во взрослом состоянии.
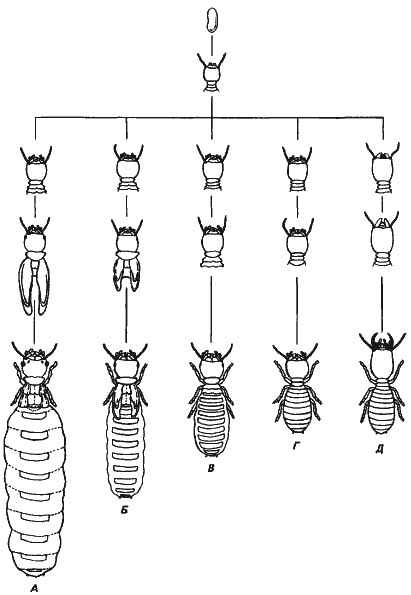
Рис. 9-9. Развитие дифференциации, приводящее к образованию различных каст у термита Amitermes atlanticus. Все касты в начале развития морфологически одинаковы, но к концу развития приобретают различную морфологию. А. Царица. Б и В. Самки-заменительницы. Г. Рабочий. Д. Солдат (Skaife, 1955; с изменениями).
Это всего лишь два из множества примеров, когда в процессе онтогенеза, направляемого единственным геномом, происходит выбор фенотипа. Потенциальные возможности, возникающие при этом для эволюционных изменений морфологии, очевидны; в гл. 6 были приведены примеры эволюции, связанной с пластичностью, которая создается переключениями развития, ведущими к различным функциональным морфологиям.
Здесь следует привести еще один пример, показывающий, что запрограммированные изменения онтогенеза можно обнаружить и у ископаемых организмов. На рис. 9-10 изображены два аммонита с гетероморфными раковинами, которые когда-то украшали собой позднемеловые моря в западной части Северной Америки. В этих раковинах запечатлена история развития животных, которые их секретировали. Обе раковины свидетельствуют о том, что это развитие делилось на три последовательных периода, причем для каждого был характерен свой особый тип роста. Вначале раковина росла по прямой, а затем в какой-то дискретный момент развития программа роста переключалась и раковина приобретала форму тортикона. Наконец, по мере приближения моллюска к зрелости направление роста вновь изменялось, что приводило к образованию терминальной U-образной жилой камеры. Помимо эстетической привлекательности раковин этих аммонитов они показывают, сколь значительной была морфогенетическая пластичность отдельного вида, допускавшая такие эволюционные изменения формы. Быстрая эволюция гетероморфов, представленная на рис. 2-4, позволяет предполагать, что в эволюции аммонитов использовалась пластичность, создаваемая переключениями с одного типа роста на другой.
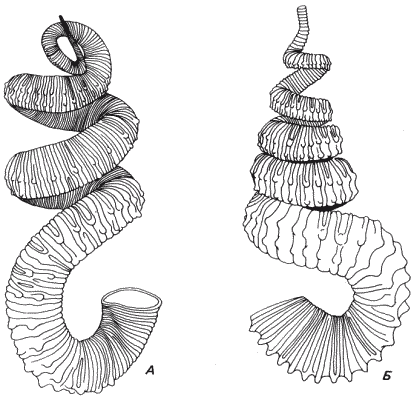
Рис. 9-10. Два гетероморфных меловых аммонита из Среднего Запада Северной Америки. У обоих видов Didymoceras nebrascense (А) и D. stephensoni (Б) для раннего периода развития раковины был характерен рост по прямой; затем его сменял период роста по спирали и, наконец, в период зрелости происходило образование последнего витка в иной плоскости (Gill, Cobban, 1973; Scott, Cobban, 1965).
В гл. 7 - 9 мы хотели продемонстрировать, что гены в самом деле контролируют онтогенез и притом весьма специфическими способами, т. е. что существует такое явление, как генетически детерминированная программа развития. Исследование этой генетической программы путем анализа мутаций, воздействующих на развитие, показало, что, хотя мутантные аллели многих генов нарушают развитие, существует небольшая группа генов, мутации которых вызывают совершенно новые эффекты. Эти гены, примером которых служат гомеозисные комплексы дрозофилы, действуют, подобно переключателям, специфицируя альтернативные типы морфогенеза. Мы подробно рассмотрели характеристики гомеозисных генов, потому что это наиболее глубоко изученные гены-переключатели, связанные с развитием. У других организмов также имеются гены-переключатели, которые, вероятно, играют важную роль в принятии решений в процессе развития, но эти гены менее изучены.
Интересная особенность генов дрозофилы, которые «принимают решения» при переключениях, связанных с детерминацией числа сегментов, их полярности и индивидуальных особенностей, состоит в том, что эти гены служат как бы для расшифровки позиционной информации. Как было показано в гл. 4, позиционная информация представляет собой важный элемент развития широкого филогенетического спектра организмов. Однако большинство беспозвоночных и позвоночных животных, служащих классическими объектами исследования в эмбриологии, слишком слабо изучены генетически, чтобы можно было расчленить генетические программы, участвующие в установлении и расшифровке пространственной организации; такую возможность предоставляет только дрозофила, чем и объясняется наша «приверженность к мухам».
Процессы развития можно анализировать классическими генетическими методами, позволяющими выявить гены, несущие регуляторные функции в живом организме. Однако при анализе генной экспрессии мы не ограничены этими методами. В сущности, при изучении большинства организмов следует пользоваться другими методами. Успехи в разработке методов клонирования генов и в тонких исследованиях ДНК и РНК дают возможность изучать гены и генную экспрессию непосредственно в процессе развития. Результаты таких работ обсуждаются в гл. 10 и 11. В гл. 12 сделана попытка свести воедино и переосмыслить материал этих и других глав, положив начало созданию эмбриогенетической основы морфологической эволюции.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Выбор места
Выбор места Для посадки клематисов необходимо учитывать некоторые особенности. Прежде всего, клематисы светолюбивы и предпочитают солнечное местоположение, но в южных районах (например, ЧПК) с жарким климатом хорошо растут и в полутени. Для посадки надо выбирать
Протонный потенциал движет бактерией
Протонный потенциал движет бактерией В 1956 году, то есть за пять лет до публикации своей знаменитой гипотезы, Митчел напечатал заметку о возможных механизмах движения флагеллярных бактерий. Один из них мы опустим за ненадобностью (он казался фантастичным и оказался
ВЫБОР МАРШРУТА
ВЫБОР МАРШРУТА Мы живем в такую эпоху, когда необходимость охраны окружающей среды овладела умами людей. Но одного желания сберечь природу недостаточно. Чтобы охранные мероприятия были действенными, чтобы в них смогло принять участие как можно большее число людей,
Гигантский потенциал
Гигантский потенциал Частота зарождения потенциальных гениев оценивается В. П. Эфроимсоном как 1/2000–1/10000, и она должна быть более-менее одинаковой у всех народностей. Эта не такая маленькая цифра. Она указывает, что по статистике как минимум в каждой второй школе страны
ВЫБОР МАРШРУТА
ВЫБОР МАРШРУТА Мы живем в такую эпоху, когда необходимость охраны окружающей среды овладела умами людей. Но одного желания сберечь природу недостаточно. Чтобы охранные мероприятия были действенными, чтобы в них смогло принять участие как можно большее число людей,
5.1. Потенциал универсальной конфигурации
5.1. Потенциал универсальной конфигурации Сопоставление ССП, сопровождающих поведение испытуемых в различных экспериментальных ситуациях, показывает, что реализации и смене поведенческого акта соответствует потенциал универсальной конфигурации (см. рис. 16.4, А).
Выбор поощрения
Выбор поощрения Используя для поощрения в дрессировке пищевую или игровую потребность собаки, особое внимание следует обращать на индивидуальный подход, в зависимости от предрасположенности к ним животных. Хотя, часто встречаются сторонники только одного из вариантов,
8.5.1. Биотический потенциал
8.5.1. Биотический потенциал Любая популяция теоретически способна к неограниченному росту численности, если ее не лимитируют факторы внешней среды. В таком гипотетическом случае скорость роста популяции будет зависеть только от величины биотического потенциала,
Выбор пищи
Выбор пищи Известно немало видов сфексов, но только три из них, насколько я знаю, встречаются во Франции. Все они любители солнца, такого горячего в области оливковых деревьев. Таковы: сфекс желтокрылый, сфекс белокаемчатый и сфекс лангедокский; все — охотники за
ВЫБОР ПИЩИ
ВЫБОР ПИЩИ Гусеница бабочки-капустницы грызет листья крестоцветных растений, к числу которых принадлежит и капуста. Шелковичный червь — гусеница тутового шелкопряда — презирает всякую растущую у нас зелень, кроме листьев тута — шелковицы. Молочайный бражник в детстве
Три эпиграфа на выбор
Три эпиграфа на выбор Под параграфом 141 в знаменитой «Риторике» М. В. Ломоносова приводится в качестве примера, «когда предыдущее с последующим противны», четверостишие Марциала, самим Михаилом Васильевичем переведённое: В тополевой тени, гуляя, муравей В прилипчивой
Можно ли изменить свой ранговый потенциал, и нужно ли это делать?
Можно ли изменить свой ранговый потенциал, и нужно ли это делать? Поскольку ранговый потенциал влияет на многие стороны жизни человека, да собственно на всю его биографию, то возникновение желания изменить его более чем ожиданно.И, понятное дело, в подавляющем
Не лучший выбор
Не лучший выбор Эта пара образуется по инстинктивным программам: Она — красавица, Он — гроза окрестных мест. Танец облегчает сближение. Обычно инициатива выбора закреплена за одним полом — на тетеревином или турухтаньем току самцы демонстрируют себя и дерутся на
Почему ранговый потенциал не растёт неограниченно?
Почему ранговый потенциал не растёт неограниченно? В самом деле, раз уж РП передаётся по наследству (пусть даже частично), а для спаривания предпочитаются ВР мужчины, то из поколения в поколение должен передаваться всё более возрастающий РП, чего не наблюдается.
Можно ли как-то скорректировать свой ранговый потенциал?
Можно ли как-то скорректировать свой ранговый потенциал? Можно. Для начала следует обратить внимание на то, что фактический РП есть функция врождённого РП и воспитующего влияния среды. В общем и среднем, фактический РП примерно на 2/3 зависит от врождённого, но это если