Роль двоичных переключателей в интерпретации позиционной информации
Роль двоичных переключателей в интерпретации позиционной информации
Возможный характер генетической реакции на позиционную информацию был описан в некотором формальном смысле Кауфманом (Kauffmann). Согласно его модели, гены, реагирующие на позиционную информацию, представляют собой набор двоичных переключателей, образующих каскады или сети. Эта модель схематически изображена на рис. 9-5. Первый ген этого пути, или каскада, дерепрессируется поступающей позиционной информацией. Эту активацию легче всего представить себе как простую реакцию на концентрацию. Если относительная концентрация морфогена достаточно высока, то переключатель приводится в соответствующее положение. Продукт первого гена в сети вызывает последующие переключения, активирующие другие гены, а продукты этих генов в свою очередь могут активировать новые локусы, что приводит к образованию каскада. Первоначальный ввод информации в систему не обязательно должен быть единичным; в модель вполне можно заложить множественные вводы, создавая избыточность и тем самым наделяя систему гомеостатическими свойствами. Если подходить к этой модели с позиций эпигенетического ландшафта Уоддингтона, то можно представить себе, что клетка проходит через ряд развилок, ведущих в долины, где на каждой развилке приводится в действие переключатель и принимается некое решение. Таким образом, на стадии выхода в каждой клетке запечатлен иерархический набор решений, связанных с выбором положений переключателей и представляющих собой уникальное отражение ее онтогенеза. Модель эта достаточно точно отражает реальное положение вещей, в чем можно убедиться на примере развития имагинального диска крыла у дрозофилы.
Гарсиа-Беллидо (Garcia-Bellido) и его сотрудники показали, что развитие диска слагается из ряда процессов компартментализации; иначе говоря, подобно тому как это происходит с полем конечности амфибии, оси данного имагинального диска определяются в результате ряда последовательных событий. Так же как и в случае поля конечности, сначала устанавливается передне-задняя ось, а за ней следуют дорсо-вентральная ось и три последовательных ограничения вдоль проксимально-дистальной оси. Каждый из образующихся при этом компартментов отделен от других, и клетки, находящиеся в одном из них, при нормальном течении развития неспособны преодолевать границы между компартментами. В крыле и груди имаго каждая клетка имеет единственное место назначения, связанное с рядом канализирующих событий, через которые она проходит в течение развития. Кауфман рассматривает эти события как отражающие ряд принятых двоичных решений. Так, клетку, находящуюся на переднем дистальном кончике крыла, следует пометить как передняя/не задняя, дорсальная/не вентральная, крыловая/не грудная, дистально-крыловая/не проксимально-крыловая. Используя модель химической волны, первоначально постулированную Тьюрингом (Turing), можно математически показать, что исходное эмбриональное поле можно последовательно подразделить серией узловых линий, положение которых зависит от первоначальной формы этого поля и от динамики роста, могущей изменить его форму или размеры. Диск крыла приближенно имеет форму эллипса. Рассмотрев особенности роста этого диска, можно предсказать местоположение узловых линий. Как показано на рис. 9-6, сходство между спроектированными на имагинальный диск действительными границами компартментов и расположением и последовательностью узловых линий очень велико. Кауфман, Шимко и Треберт (Kauffman, Shymko, Trabert) провели сходное сопоставление, спроектировав узловые линии на зародыш, находящийся на стадии клеточной бластодермы. В этом случае различные участки зародыша специфицируются в результате ряда последовательных двоичных решений. Как схематически показано на рис. 9-7, эта модель предсказывает, что зародыш делится поперек сначала надвое, затем на четыре части, далее на восемь и наконец делится в дорсо-вентральной плоскости. В пределах каждой ограниченной таким образом области клеткам можно приписать двоичный адрес при помощи четырех гипотетических переключателей, которые могут находиться в состоянии либо 0, либо 1. Так, первый переключатель находится в состоянии 1 в передней половине и в состоянии 0 - в задней. В следующем наборе зон, определяемых узловыми линиями, второй переключатель находится в состоянии 0 в передних и задних четвертях и в состоянии 1 - в двух центральных четвертях. В результате продолжения такого комбинаторного кодирования каждая область получает свое особое обозначение.
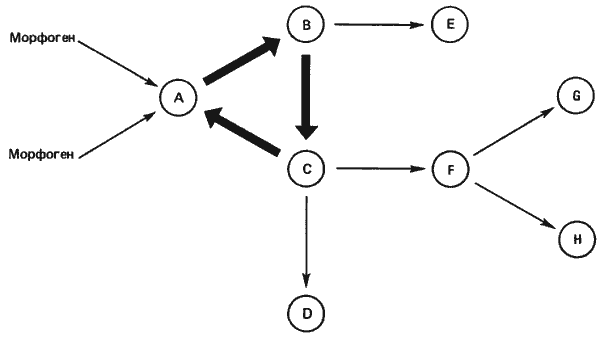
Рис. 9-5. Схема гипотетического набора генных переключателей, образующих замкнутый контур. Внешний индуктор или индукторы (морфогены) активируют ген А, который в свою очередь активирует ген В, ген В - ген С, а ген С - снова ген А, так что образуется замкнутый контур. Затем внешний индуктор можно удалить, но система остается при этом «включенной». В данной системе гены В и С действуют также и за пределами контура, активируя другие локусы (Kauffman, 1972; с изменениями).
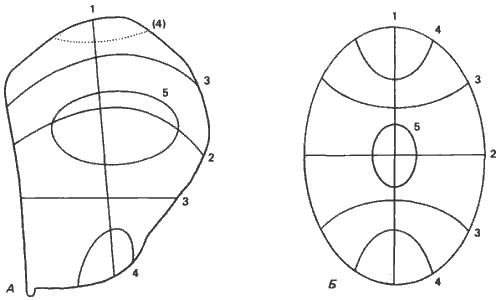
Рис. 9-6. Схема реальных и теоретических пограничных линий между компартментами в имагинальном диске крыла Drosophila melanogaster. А. Линии наложены на карту презумптивных зачатков диска. Цифры возле каждой линии указывают на последовательность, в которой, согласно наблюдениям, границы между компартментами ограничивают потенции клеток в этом диске (пунктирная линия 4 на самом деле не наблюдалась). Б. Линии, предсказанные моделью Тьюринга для эллипса. Сходство между двумя схемами поразительно (Kauffman, Shymko. Trabert, 1978; с изменениями).
Аналогичная система обозначений была привлечена для того, чтобы объяснить, каким образом детерминируется индивидуальность каждой имагинальной структуры, образующейся в различных сегментах. Так, различным имагинальным дискам можно дать обозначения, используя коды с комплектом из пяти переключателей. Конечно, этой модели внутренне присуща известная иерархия, причем некоторые сегменты и диски в этой иерархии более тесно связаны между собой, чем другие. Этот аспект данной модели хорошо согласуется с замечаниями Хадорна (Hadorn) относительно трансдетерминационных событий между имагинальными дисками: когда фрагменты дисков, выращиваемых в культуре, переходят из одного детерминированного состояния в другое, это происходит совсем случайным образом. Как и в комбинаторном коде, при этом выявляется некая иерархия. Кроме того, некоторые трансдетерминации никогда не происходят, например диск крыла никогда не превращается в хоботок. Кауфман объясняет это тем, что в дифференцировке этих двух структур участвуют два набора переключателей, и для такого превращения оба они должны вступить в действие, что представляется в высшей степени маловероятным. Помимо того что комбинаторный код позволяет объяснить трансдетерминационные взаимоотношения, он согласуется также с гомеозисными превращениями, наблюдаемыми у дрозофилы. Используя кодовые обозначения, примененные для различных областей бластодермы (рис. 9-7), и наложив на эту карту положение имагинальных дисков, развившимся из них имагинальным структурам можно дать обозначения в соответствии с комбинаторным кодом.
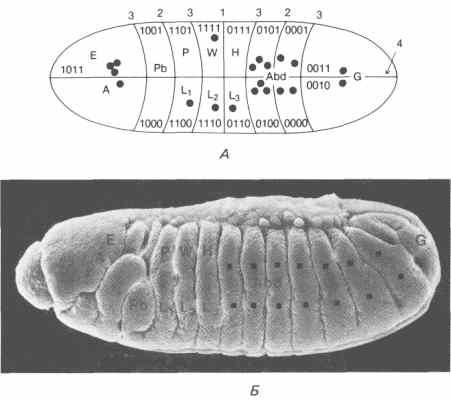
Рис. 9-7. А. Ряд пограничных линий, разделяющих компартменты, наложенные на зародыш дрозофилы на стадии клеточной бластодермы. Каждый компартмент несет предложенные Кауфманом метки по двоичному коду, специфицирующие индивидуальность каждого сегмента. ? - глаз; А - антенна; Pb - хоботок; Р - переднегрудь; L1, L2 и L3 - 1-я, 2-я и 3-я ноги; W - крыло; Н - жужжалъце; Abd - брюшко; G - половые органы (Kauffman, Shymko, Trabert, 1978). Б. Микрофотография сегментированного зародыша дрозофилы, сделанная с помощью сканирующего электронного микроскопа. Обозначения на микрофотографии указывают сегменты, из которых, как предполагается, развиваются имагинальные структуры, указанные на рис. А. (Фотография любезно предоставлена д-ром F. R. Turner.)
Большую часть гомеозисных превращений, наблюдаемых среди этих структур, можно объяснить изменениями в одном переключателе. Так, для того чтобы антенна (1010) превратилась во вторую ногу (1110), достаточно одного переключения 0 —> 1. Это, в сущности, и проделывает мутация Antennapedia, и Кауфман высказывает мнение, что некоторые гомеозисные локусы, возможно, представляют собой гены-переключатели, необходимые для расшифровки позиционной информации. Переключатели эти обладают еще одним свойством, которое также исследовал Кауфман. После того как ген включен или активирован, он остается в таком состоянии. Гены, очевидно, обладают памятью. Предполагается, что эта память существует в форме ряда «замкнутых контуров». Согласно этой модели, после того как под действием какого-либо внешнего сигнала (например, позиционной информации) первый ген контура приходит в активное состояние, он активирует второй локус, который в свою очередь активирует третий. Как второй, так и третий локусы сами могут функционировать как активаторы, дерепрессирующие другие локусы. Как показано на рис. 9-5, в этом простом варианте одним из локусов, дерепрессируемых третьим геном, является первый ген. В таком случае ген 3 участвует в создании петли обратной связи, которая будет усиливать первоначальную активацию гена 1. Кроме того, можно представить себе, что гены 2 и 3 сами поддерживаются в активном состоянии другими локусами, еще больше усиливая действие контура. Простым конкретным примером системы этого типа служит лактозный оперон Escherichia coli. Снабжая клетки Е. coli лактозой, можно индуцировать у них синтез фермента ?-галактозидазы. Однако, для того чтобы стала возможной индукция, в клетку должно проникнуть некоторое количество субстрата. Это происходит следующим образом. Вначале в бактерию просто просачивается лактоза, индуцируя синтез соответствующего катаболического фермента и пермеазы. Синтезированная пермеаза включается в плазматическую мембрану клетки, облегчая перенос в клетку лактозы. Первоначальная активация данной системы зависит, таким образом, от проникновения в нее лактозы, что в свою очередь зависит от концентрации. При субоптимальных концентрациях синтез ферментов не индуцируется и клетки не смогут использовать лактозу в качестве источника углерода. Если популяцию Е. coli выращивать при такой концентрации лактозы, которая чуть выше индуцирующего уровня, то клетки постепенно приобретают способность использовать этот сахар. При этом одни клетки индуцируются, а другие нет. Если, в то время когда примерно половина составляющих популяцию клеток индуцирована, концентрацию лактозы понизить путем разведения до субоптимального уровня, то клетки, которые уже были индуцированы, остаются в этом состоянии, а неиндуцированные остаются неиндуцированными. Эти свои состояния клетки сохраняют даже после деления, т.е. дочерние клетки «помнят». Такую «память» обеспечивает пермеаза, включающаяся в мембраны индуцированных клеток. Пермеаза облегчает перенос лактозы через клеточные мембраны, даже если концентрация ее невелика. Это в свою очередь индуцирует дальнейшее образование пермеазы, усиливающей состояние индуцированности. Следовательно, составляющие популяцию клетки находятся в двух альтернативных состояниях - активированном и неактивированном. Однако, для того чтобы приблизить эту ситуацию к гипотетической ситуации Кауфмана, необходимо наличие внутреннего индуктора для лактозного оперона, который поддерживал бы его активность даже в отсутствие внешнего индуктора. Как и в случае теории позиционной информации, эти идеи о генных переключателях и комбинаторном кодировании создают теоретическую основу, в рамках которой можно рассматривать генетическую регуляцию развития. На самом деле недавние данные о генетической регуляции основного типа сегментации у зародыша дрозофилы показали, что в деталях модель комбинаторного кодирования неверна; однако идея о последовательной компартментализации вполне оправданна.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ДОСТУП К ИНФОРМАЦИИ
ДОСТУП К ИНФОРМАЦИИ Жизнь животных тесным образом связана со средой, в которой они обитают. Среда обеспечивает их пищей, водой, кислородом, убежищем. Животные постоянно должны опасаться врагов и искать контакта с родственными организмами, хотя бы для того, чтобы
1.1. Понятие генетической информации
1.1. Понятие генетической информации Как хранится и воспроизводится генетическая информация? Какие структуры организма являются ее «хранилищем»? Как генетическая информация проявляется и как изменяется в ряду поколений? Без понимания этих вопросов немыслимо подходить
1.3. Проявление генетической информации
1.3. Проявление генетической информации Как проявляется генетическая информация, локализованная в ДНК-содержащих структурах? Ее проявление отражается в фенотипе, который, согласно современным представлениям, всегда является результатом взаимодействия генов в
ДОСТУП К ИНФОРМАЦИИ
ДОСТУП К ИНФОРМАЦИИ Жизнь животных тесным образом связана со средой, в которой они обитают. Среда обеспечивает их пищей, водой, кислородом, убежищем. Животные постоянно должны опасаться врагов и искать контакта с родственными организмами, хотя бы для того, чтобы
Полезные мутации переключателей
Полезные мутации переключателей Устойчивость к ядам, вирусам, бактериям и прочим паразитам, защитная окраска, превращение опадающих семян в неопадающие — все это примеры простых адаптаций, для развития которых бывает достаточно одной-двух удачных мутаций, поддержанных
3. КОДИРОВАНИЕ ИНФОРМАЦИИ
3. КОДИРОВАНИЕ ИНФОРМАЦИИ Кодированием называют совершаемое по определённым правилам преобразование информации в условную форму – код. В сенсорной системе сигналы кодируются двоичным кодом, т.е. наличием или отсутствием электрического импульса в тот или иной момент
Отсутствие образования и информации
Отсутствие образования и информации Чтобы учить других, необходимо, прежде всего, чтобы врач сам обладал достаточной информацией. Думаю, что нет необходимости утверждать, что во время обучения в университете будущий врач не получает по данной проблеме абсолютно никакой
3.3. Свойства генетической информации
3.3. Свойства генетической информации Одним из основателей современной генетики, выдающимся датским ученым В. Иоганнсеном были предложены базовые генетические термины: ген, аллель, генотип, фенотип. Введение термина «аллель» позволяет определить генотип как
Проявление генетической информации
Проявление генетической информации Как же взаимосвязаны между собой генотип и фенотип? Или, другими словами: как проявляется генетическая информация? Согласно современным представлениям, генотип – это система взаимодействующих генов, а фенотип – это результат
О ДРУГИХ ИСТОЧНИКАХ ИНФОРМАЦИИ
О ДРУГИХ ИСТОЧНИКАХ ИНФОРМАЦИИ Круг вопросов, рассмотренных в этой книге в отечественной литературе освещается, пожалуй, впервые и уже поэтому работа не может претендовать на полноту и разрешение всех, возникающих у владельцев проблем. Автор надеется, что эта первая
Проблема с теорией информации
Проблема с теорией информации Создание теории информации было очень важным достижением. Оно позволило нам понять, как физическое явление – электрический импульс – может стать психическим явлением – нервным сигналом (сообщением). Но с первоначальной формулировкой была
О записи информации
О записи информации Вот поэтому аналогию себе нам надо искать в записях информации без участия нас самих — в чем-то таком, что запоминает информацию само, без нашей воли, образно говоря, в том, что находится у нас «на поводке».Пример. Вот, скажем, какой-то музыкальный
Гравитация как носитель информации
Гравитация как носитель информации При обсуждении одной из тем этой книги нетерпеливый комментатор выразил неудовольствие, которое, думаю, могут выразить и другие читатели: «Читаю-читаю, жду-жду, когда же вы все-таки к результату-то придете, а вы все кругами да по