Глава 9 Становление пространственной организации
Глава 9
Становление пространственной организации
Пространственная организация и происхождение формы
Главная проблема, стоящая перед биологами, занимающимися изучением развития, - объяснить механику процессов, в результате которых из одноклеточной зиготы образуется морфологически более сложный многоклеточный взрослый организм. С генетической точки зрения это влечет за собой необходимость объяснить, каким образом закодированная в ДНК одномерная информация реализуется в трехмерной структуре организма. Программа развития слагается из связанных между собой явлений двух типов - клеточной дифференцировки и становления пространственной структуры. Возможность отделить друг от друга эти два аспекта онтогенеза нетрудно продемонстрировать на довольно простом примере. Если произвести биохимический анализ правой и левой рук человека и перечислить все образующие их мышцы, сухожилия, кости и т. п., то они окажутся идентичными. Между тем одного взгляда на эти два органа достаточно, чтобы убедиться в том, что они не идентичны. Одна рука не может заменить другую. Еще яснее это видно при сравнении руки и ноги. В настоящей главе мы и попытаемся выяснить, каким образом возникают эти различия, т.е. как происходит развитие пространственной структуры и формы. В конечном счете эволюцию и морфологии, и клеточной дифференцировки следует понимать именно в контексте становления пространственной структуры.
Один подход к изучению процесса онтогенеза был выдвинут в 40-х годах Уоддигтоном (Waddington). В процессе своего развития клетка проходит то, что Уоддингтон назвал эпигенетическим ландшафтом. Эпигенетический ландшафт (рис. 9-1) представляет собой равнину, изрезанную рядом долин. Долины берут начало на возвышенном конце равнины и тянутся вниз; при этом они постепенно расходятся, заканчиваясь каждая в своей особой точке на нижнем конце равнины. Клетка движется от верхнего конца равнины к нижнему по системе долин. В каждой точке ветвления клетка должна принять некое морфогенетические решение, в результате чего ее потенции к развитию ограничиваются. Находясь на верхнем конце ландшафта, клетка теоретически может достигнуть каждой из особых конечных точек. Однако, после того как она примет свое первое решение, например в первой точке ветвления, ей останется доступным только одно из подмножеств конечных точек. Клетка начинает свой путь в состоянии тотипотентности, но постепенно ее возможности становятся все более и более ограниченными в результате принимаемых ею решений. Уоддингтон назвал этот процесс канализацией. По своим общим свойствам процесс канализации приложим как к клеточной дифференцировке, так и к становлению пространственной структуры. Точки принятия решений - развилки долин - подвержены воздействиям внешних сил, например гормональных стимулов или индукции, а решение зависит от обусловленной генетически реакции клетки на данный стимул. Как станет ясно из дальнейшего, наблюдаемые нами решения, которые клетка принимает в процессе онтогенеза, относятся к типу «или-или», и их можно рассматривать как ряд двоичных решений типа «направо-налево». Расстояние от верхнего конца ландшафта к нижнему соответствует времени развития. Эту модель можно использовать как для мозаичного, так и для регуляционного развития, просто перемещая точки ветвления долин либо к верхнему, либо к нижнему концу холма соответственно. Хотя данная модель в том виде, в каком она здесь описана, позволяет представить себе процесс развития, она не объясняет его. Кроме того, эта модель статична, поскольку в ней формально не представлен процесс клеточного деления. Тем не менее идея о ряде канализирующих событий очень важна и отражается, например, в развитии конечности позвоночных, в частности в детерминации осей, определяющих отличие правой конечности от левой.
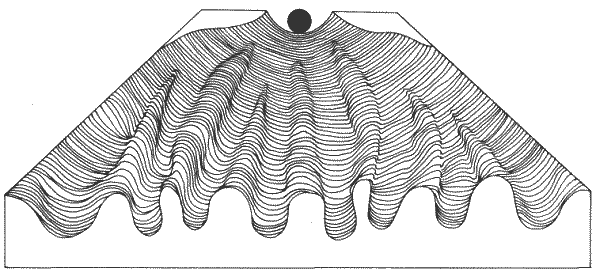
Рис. 9-1. Эпигенетический ландшафт Уоддингтона. Шарик на вершине изображает клетку, а долины под ним - различные пути развития, по которым она может пойти (Waddington, 1966).
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава I. Принципы работы и организации мозга
Глава I. Принципы работы и организации мозга Поведение животных складывается из добывания пищи, размножения, межвидовых и внутривидовых взаимодействий. Животные едят, размножаются, мигрируют, приспосабливаются к изменчивым условиям среды, ухаживают за потомством,
2.1. Становление и основные положения синтетической теории эволюции
2.1. Становление и основные положения синтетической теории эволюции Эволюционизм возник как альтернатива учению о неизменности видов. Вопросы, связанные с возникновением и развитием жизни, прошли через всю интеллектуальную историю человечества. Количество литературы,
3.2. Становление этологии и вопросы эволюции поведения
3.2. Становление этологии и вопросы эволюции поведения Этология возникла в середине 1930-х гг. как наука, изучающая поведение животных в естественной среде обитания. Эта наука дала миру целую плеяду талантливых ученых, однако даже на таком ярком фоне выделяются имена ее
4.1. Типы социальной организации
4.1. Типы социальной организации В этологических исследованиях был обнаружен широкий диапазон различных видов социальной организации. У высших животных их социальные структуры и социальное поведение в процессе эволюции достигают исключительной степени сложности и
10.1. Становление взглядов на проблему когнитивных способностей животных
10.1. Становление взглядов на проблему когнитивных способностей животных Первые систематизированные данные по исследованию когнитивных способностей животных получил английский ученый, ученик Ч. Дарвина Дж. Романес (1848–1894). В 1883 г. выходит его книга «Интеллект животных».
11.1. Становление психологии
11.1. Становление психологии В дословном переводе «психология» означает «наука о душе». В научной терминологии вместо слова «душа» стали употреблять слово «психика», что, однако, не прибавило определенности этому понятию. Сложность, неопределенность, «неуловимость»
Становление локализации и пространственной организации
Становление локализации и пространственной организации Во время оогенеза происходит чрезвычайно активная транскрипция генов и накопление ооцитами мРНК. Накапливающиеся в ооцитах мРНК столь разнообразны по своим последовательностям, что, как только после начала
Становление пространственной организации и спецификация местоположения
Становление пространственной организации и спецификация местоположения Презумптивную область ноги, которую пересаживал Гаррисон, можно рассматривать как эмбриональное поле. В более общем смысле поле - это участок зародыша или группа клеток, в пределах которых может
Становление пространственной организации и полярные координаты
Становление пространственной организации и полярные координаты Хадорн (Hadorn) и его ученики обнаружили, что если у личинки третьего возраста дрозофилы удалить один из имагинальных дисков и пересадить его путем инъекции новой личинке-реципиенту, то он будет
Глава 5. УНИВЕРСАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ БЛОКИ КАК ОСНОВА ОРГАНИЗАЦИИ СЛОЖНЫХ ФУНКЦИЙ
Глава 5. УНИВЕРСАЛЬНЫЕ ФУНКЦИОНАЛЬНЫЕ БЛОКИ КАК ОСНОВА ОРГАНИЗАЦИИ СЛОЖНЫХ ФУНКЦИЙ Анализ процессов пищеварения и транспорта на современном уровне и в особенности эволюции этих процессов открывает новые горизонты для понимания принципов организации сложных
4.2. Возникновение клеточной организации
4.2. Возникновение клеточной организации Учеными было доказано, что после испарения воды из реакционного объема в амфифильных липидоподобных и липидных молекулах формируются жидкокристаллические агрегаты, в которых молекулы расположены периодическими слоями, как в
Становление ученого
Становление ученого Его жизнь прошла в беспрерывной борьбе и беспримерных дерзаниях. Десятилетиями длилась эта борьба. Трудно было поверить, что он обнаружит так много сил, найдет в себе столько решимости. Безудержная мысль Александра Васильевича Вишневского и на