Гомеозис и гомеозисные мутации
Гомеозис и гомеозисные мутации
Действие генов теснейшим образом связано с онтогенезом, и эта их связь выявляется при возникновении мутаций, которые резко прерывают развитие организма. Существуют, однако, мутации другого класса, которые изменяют процесс онтогенеза, но не прерывают его. Это гомеозисные мутации. На важную роль и теоретическое значение этого рода изменения развития впервые указал Уильям Бэтсон (William Beateson) в своей книге «Материалы к изучению изменчивости», опубликованной в 1894 г. Его соображения при создании термина «гомеозис» и определение этого термина все еще сохраняют силу и привлекают внимание к наиболее существенным чертам этой концепции.
«Превращение антенны насекомого в ногу, глаза ракообразного - в антенну, лепестка - в тычинку и тому подобное - все это примеры одного рода. Желательно и даже необходимо, чтобы такие изменения, состоящие в том, что один из членов ряда меристических структур принимает форму или приобретает признаки, свойственные другим членам ряда, были выделены в особую группу явлений ... Поэтому я предлагаю ... термин гомеозис , ..., поскольку главное здесь не в том, что произошло некое изменение, а в том, что одно, изменившись, приобрело сходство с чем-то другим».
Затем Бэтсон переходит к перечислению примеров гомеозисных изменений у столь различных организмов, как млекопитающие и кольчатые черви. Если говорить о млекопитающих, то описано, например, несколько редких особей вымерших неполнозубых, у которых в крестцовом отделе позвоночника обнаружены позвонки, похожие на грудные. Гораздо чаще, однако, гомеозисные изменения встречаются у членистоногих - животных, целиком построенных из ряда метамерных сегментов; а из членистоногих больше всего сведений о типах и механизмах гомеозиса получено на насекомых.
Остатки некоторых из древнейших вымерших насекомых найдены в слоях, относящихся к каменноугольному периоду. У этих насекомых, как и у современных крылатых насекомых, было четыре крыла, сходные по морфологии с крыльями ныне живущих видов. В отличие от современных насекомых у них была, кроме того, пара крыловидных придатков, или паранотальных лопастей, отходящих от спинки первого грудного сегмента. Эти лопасти считались возможным свидетельством того, что крылья возникли как выступающие наружу складки интегумента. Они могли первоначально служить органами, помогающими насекомому планировать. Примитивные признаки - расположение крыльев на втором и третьем грудных сегментах и наличие паранотальных лопастей - у современных насекомых отсутствуют. Однако они могут возникать в результате гомеозисной мутации у таракана Blattella germanica. Росс (Ross) описал у этого примитивного насекомого наследственную мутацию Prowings, вызывающую развитие крылоподобных придатков на спинке переднегруди. Сходное гомеозисное изменение описал Гершкович (Herskowitz) у Drosophila melanogaster.
Другая сцепленная с полом рецессивная мутация labiopedia описана у хрущака Tribolium confusum. У особей, гомо- или гемизиготных по этой мутации, лабиальные щупики превращаются в грудные ноги. Такое превращение наблюдается как на личиночной, так и на взрослой стадии (рис. 8-1). При этом происходит полная перестройка, вплоть до развития мускулатуры, обычно имеющейся у ног. Однако эти мышцы, очевидно, лишены иннервации, так как лабиальные ноги неподвижны.
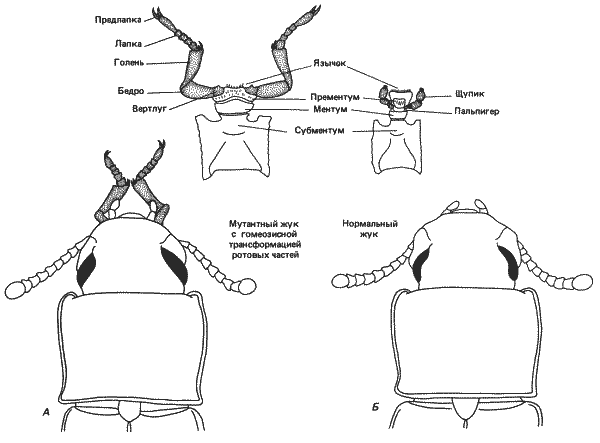
Рис. 8-1. Головы нормальной (Б) и мутантной - labiopedia, (А) особей хрущака Tribolium confusum со спинной стороны. Показаны элементы ротовых частей, превращающиеся у мутантных особей в конечность. Дистальные части нормальных губных элементов (щупик и пальпигер) превращаются в обычную ногу с вертлугом на проксимальном конце и тарзальными коготками на дистальных концах (Daly, Sokoloff, 1965).
Более резко выраженные трансформации наблюдаются у тутового шелкопряда Bombyx mori (чешуекрылые). У этого насекомого имеется комплекс мутаций, известных под названием серии E-аллелей (Extra legs). Его гусеница обладает очень своеобразной морфологией: за темно окрашенной головой с находящимися на ней глазами следуют три грудных сегмента, несущие членистые ножки. На спинной стороне второго грудного сегмента имеется темное глазчатое пятно. Брюшко состоит из восьми сегментов, за которыми следует концевой хвостовой сегмент. Брюшные сегменты 3-6 и хвостовой сегмент снабжены мясистыми ходильными ножками. На спинной поверхности второго брюшного сегмента имеется пигментное пятно в форме полумесяца, а на спинной поверхности пятого - пятно в форме звезды. Морфология гусеницы дикого типа показана на рис. 8-2, А. Доминантные мутантные аллели Е-серии вызывают перемещение этих пятен на другие сегменты. Так, у особей, гетерозиготных по мутации EEl (Extra Crescents and legs), голова и грудь такие же, как у дикого типа, но в отличие от последних на первом и втором брюшном сегментах у них имеются ходильные ножки, а на третьем грудном и первом брюшном - пигментные пятна в форме полумесяца. Мутация EN (Extra legs - New additional crescent) вызывает появление полумесяца на третьем и звезд - на шестом, седьмом и восьмом брюшных сегментах; на пятом брюшном сегменте звезда исчезает. Эти модификации можно объяснить как результаты изменения индивидуальностей (специфических особенностей) брюшных и грудных сегментов (как на спинной, так и на брюшной их сторонах). В наиболее резкой форме эти изменения проявляются у гомозигот по упомянутому выше аллелю EN. Такой генотип детален и приводит к гибели у личинок ранних личиночных возрастов. Морфология особей EN/EN показана на рис. 8-2, Г. Все брюшные сегменты превращены у них в грудные сегменты, снабженные членистыми конечностями, характерными для этих метамеров.
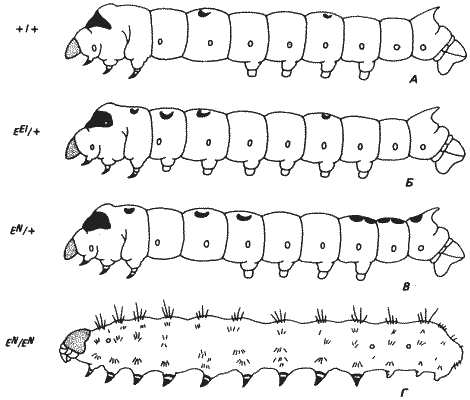
Рис. 8-2. Нормальная и мутантная личинки тутового шелкопряда Bombyx mori. А. Нормальная личинка пятого возраста: от первых трех сегментов отходят пигментированные ходильные ноги; на дорсальной поверхности второго и пятого брюшных сегментов имеются пигментированные пятна, а от вентральной поверхности брюшных сегментов 3-6 отходят мясистые ходильные ноги. Б. Личинка пятого возраста, гетерозиготная по доминантной мутации ЕEl (Extra crescents and legs). Обратите внимание на добавочные серповидные пятна на третьем грудном и первом брюшном сегментах и на наличие ходильных ног на двух первых брюшных сегментах. Создается впечатление, что два первых брюшных сегмента и третий грудной сегмент приобрели некоторые признаки задних брюшных сегментов. В. Личинка пятого возраста, гетерозиготная по доминантной мутации EN (Extra legs new additional crescent), у которой наблюдаются сходные изменения пигментации на дорсальной стороне тела, но нет изменений на вентральной стороне. Г. Поздний зародыш, гомозиготный по аллелю EN. Обратите внимание на наличие грудных ног на большей части брюшных сегментов; у нормальных особей такие придатки имеются только на первых трех сегментах тела (грудных) (Tanaka, 1953).
Гомеозисные мутации, аналогичные наблюдаемым у жуков и бабочек, обнаружены также у двукрылых - насекомых, достигших наиболее высокого развития. Мутации ВХ-С (Bithorax Complex) Drosophila melanogaster приводят к таким же изменениям сегментов, как и мутации Е-аллелей. У взрослых особей дрозофилы, гомозиготных по двум рецессивным мутациям, bx (bithorax) и pbx (postbithorax), обнаружены удивительные изменения, показанные на рис. 8-3. Подобно всем двукрылым, дрозофила обладает лишь одной парой крыльев, отходящих от второго грудного сегмента взрослой особи. Вторая пара крыльев, располагающаяся у других крылатых насекомых на третьем грудном сегменте, у них редуцировалась до маленьких утолщений, называемых жужжальцами или органами равновесия. У двойных мутантов bx pbx эти жужжальца превращаются в крылья. Кроме того, у взрослой мутантной особи третий грудной сегмент, который у дрозофилы обычно бывает довольно маленьким, становится похожим на большой второй грудной сегмент.
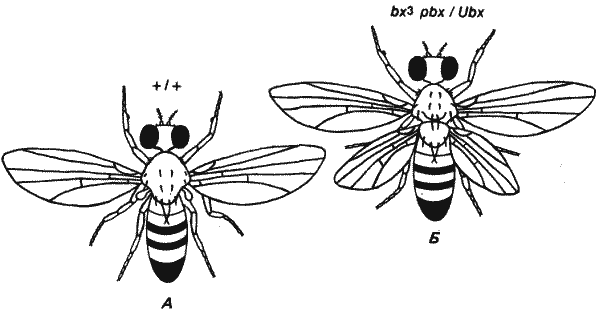
Рис. 8-3. Нормальная (А) и мутантная (Б) взрослые особи дрозофилы. Гомеозисные мутации (bx3pbx/Ubx) вызьшают превращение обычно небольшой спинки заднегруди и жужжалец в хорошо заметную среднегрудь и вторую пару крыльев.
У дрозофилы встречается также ряд мутаций, превращающих ротовые части в другие структуры. Эти гены составляют другой генный комплекс ANT-C (Antennapedia Complex). При мутации pb (proboscipedia) у дрозофилы, так же как и при мутации labiopedia у хрущака, лабиальные щупики превращаются в ходильные ноги. Однако некоторые аллели proboscipedia вызывают превращение щупиков в антенны (рис. 8-4). Другая доминантная мутация Antp (Antennapedia) вызывает превращение, которое упоминается в первоначальном бэтсоновском определении гомеозиса. При этом антенны у взрослой мухи превращаются в полностью сформированные ноги, отходящие от головы мухи в стороны (рис. 8-4). Задача, конечно, состоит в том, чтобы, не ограничиваясь простым перечислением этих дефектов, попытаться объяснить, как возникают эти кажущиеся магическими изменения.
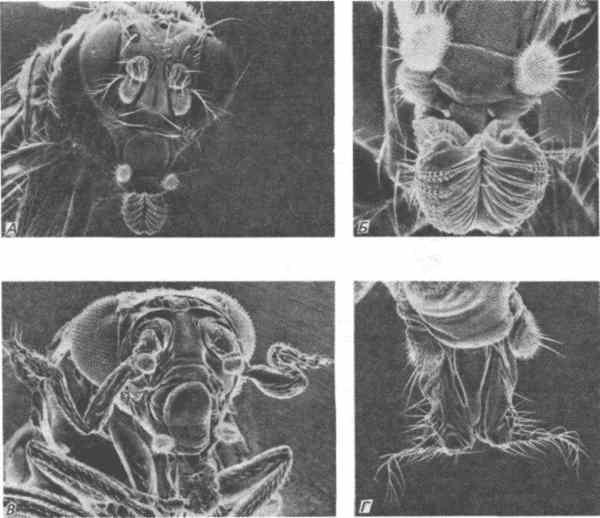
Рис. 8-4. Микрофотографии головы и ротовых частей нормальных и мутантных особей Drosophila melanogaster, сделанные при помощи сканирующего электронного микроскопа. А. Голова нормальной особи. Б. Ротовые части нормальной особи. В. Особь, гетерозиготная по мутации Antennapedia, у которой в результате гомеозисного превращения члеников антенн на голове образовались ноги, содержащие большую часть члеников нормальной ноги; однако дистальные элементы этой структуры все еще сохраняют сходство с концевыми члениками антенны. Г. Ротовые части особи, гомозиготной по мутантному аллелю proboscipedia. Обычные псевдотрахеальные ряды микрощетинок нижней губы замещены структурами, напоминающими аристы, или концевые членики нормальной антенны. Другие мутантные аллели в этом локусе вызывают превращение нижнегубных лопастей в ноги. (Фотографии любезно предоставлены д-ром Turner.)
Самые обширные генетические исследования по гомеозисным дефектам проводятся на Drosophila melanogaster. У этого насекомого имеется широкий набор мутаций, вызывающих более или менее значительные изменения характера сегментов, и ряд работ, в которых изучались влияния этих мутаций на развитие, помогли выявить генетические механизмы, участвующие в определении общего плана строения тела и индивидуальности отдельных сегментов, поскольку они регулируются гомеозисными локусами. В целом экспрессия класса гомеозисных мутаций заключается в изменении направления детерминации. Поэтому их считали мутациями, нарушающими интерпретацию позиционной информации, определяемой материнским геномом в яйце и раннем зародыше. Кроме того, их рассматривали как морфогенетические двоичные переключатели, транслирующие позиционную информацию по типу «или-или». Однако группа гомеозисных мутаций довольно неоднородна как по своим генетическим свойствам, так и по своему влиянию на развитие. Распределение гомеозисных локусов, идентифицированных у Drosophila melanogaster, по ее четырем хромосомам схематически показано на рис. 8-5. Гомеозисные локусы имеются во всех ее хромосомах и содержат как доминантные, так и рецессивные аллели.
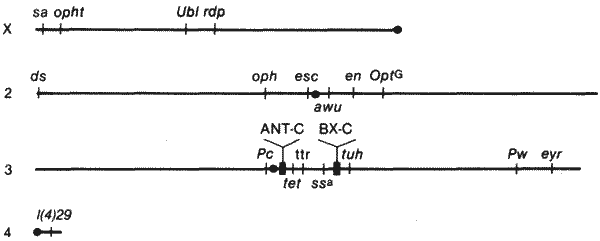
Рис. 8-5. Схема распределения большинства известных гомеозисных мутаций между четырьмя хромосомами Drosophila melanogaster. Черными кружками показано положение центромеры в Х-хромосоме и во 2-й, 3-й и 4-й хромосомах. На правом плече 3-й хромосомы имеются два кластера гомеозисных локусов: ANTC-C (Antennapedia) и BX-C (bithorax). Остальные сокращения означают (в порядке их расположения на карте): sa - sparse arista; opht - ophtalmoptera; rdp - reduplicated; Ubl - Ultrabithoraxlike; ds - dachsous; oph - ophtalmopedia; esc - extra sex comb; awu - augenwulst; en - engarailed; OptG - Ophtalmoptera of Goldschmidt; Pc - Polycomb; tet - tetraltera; ssa - spineless aristapedia; tuh - tumorous head; Pw - Pointed wing; eyr - eyes-reduced; 1(4)29 - lethal 29 (в 4-й хромосоме).
Разнообразие свойств, наблюдаемых при этих генетических дефектах, заставляет думать, что они, возможно, различаются по механизмам, участвующим в реализации каждого отдельного превращения. Иными словами, некоторые из них могут быть связаны с генами-переключателями, непосредственно участвующими в процессах детерминации, тогда как другие не связаны с ними. На это указывает тот факт, что мутантные аллели нескольких из этих локусов обладают плейотропным действием, вызывая не только гомеозисные, но и другие неспецифические аномалии фенотипа. Хорошим примером служит мутация ssa (spineless-aristapedia), которая вызывает превращение антенны в ногу, сходное с описанным для доминантной и неаллельной к ней мутации Antp. Помимо этих гомеозисных превращений мутанты ssa отличаются опущенными крыльями, тонкими щетинками и слиянием члеников лапки; таких эффектов нельзя было ожидать от гена-переключателя, определяющего выбор между двумя альтернативными типами развития. Поэтому мы можем сделать вывод, что аллель ssa детерминирует гомеозис косвенным путем, быть может, через какой-то дефект, затрагивающий метаболизм. Возможную природу этого косвенного воздействия вскрывает анализ группы диск-специфичных летальных мутаций дрозофилы, проведенный Ширном (Shearn) и его сотрудниками. Эти нарушения не оказывают прямого действия на эмбриогенез в той мере, в какой это относится к развитию нормальной личинки. Однако имагинальные диски, из которых при нормальном развитии во время метаморфоза образуется интегумент взрослой особи, у мутантов не пролиферируют или же вообще не формируются. Обнаружены температурочувствительные аллели для нескольких из этих локусов. Перенос мутантных личинок в условия непермиссивной температуры в разные сроки на стадии пролиферации имагинальных дисков приводит не к элиминации соответствующих дисков, а к гомеозисным трансформациям у взрослых особей, получающихся из этих личинок. Поскольку было показано, что ко времени изменения температуры диски уже были детерминированы, мутация, очевидно, вызывает изменение детерминации после ее первоначального возникновения. Такого же рода изменение наблюдали Е. Хадорн (Е. Hadorn) и его ученики при выращивании имагинальных дисков in vitro. Диски, эксплантированные в брюшную полость взрослых самок, растут, а если разрезать их на части, то у них продолжается клеточное деление. Если ввести такие фрагменты в метаморфизирующих личинок, то они сами метаморфизируют. В большинстве случаев из такого выращенного диска образуется структура, соответствующая диску, первоначально взятому в культуру; т.е. из диска крыла получаются структуры крыла, даже если он выращивался в культуре в течение нескольких лет. Именно такого рода эксперименты позволили показать, что детерминация ткани диска происходит рано и ее дифференцировка завершается на стадии куколки. Однако из полученных таким образом дисков после метаморфоза не всегда развиваются ожидаемые структуры, а иногда получаются производные других дисков. Это явление было названо трансдетерминацией. Такое изменение детерминации вызывается у этих клеток не единичным мутационным событием, как в случае гомеозиса, а, скорее, изменением характера генной экспрессии. Дальнейшее изучение этого явления, проведенное Стребом (Strub), показало, что не все клетки данного диска (в частности, диска ноги) способны к трансдетерминации. Для того чтобы данная группа клеток могла реализовать свою потенциальную способность к трансдетерминации, она должна находиться по соседству с разрезанным или повержденным участком диска и в ней должно происходить деление клеток. Возможно, что мутации discless, изучавшиеся Ширном, и мутации ssa вызывают гомеозисные нарушения, стимулируя эти события (т.е. локальные повреждения клеток или их гибель, за которыми следует компенсаторная пролиферация за счет соседних клеток) in situ. Таким образом, изменение судьбы клеток вызывается изменением направления их детерминации, как это происходит при трансдетерминации, а не воздействием на первоначальное детерминирующее событие.
Другую косвенную причину гомеозиса можно продемонстрировать на примере мутаций в локусе Ubl (Ultrabithoraxlike), упоминавшихся в гл. 7. Мортен и Лефевр (Mortin, Lefevre) показали, что эта сцепленная с полом доминантная мутация имитирует аутосомную мутацию Ubx (Ultrabitorax), которая близка к упоминавшимся выше мутациям bx и pbx. Мутация Ubx, подобно bx и pbx, вызывает превращение жужжалец взрослых особей в крылья и некомплементарна ни мутации bx, ни мутации pbx (см. рис. 8-3). Мутация Ubl вызывает аналогичную трансформацию и взаимодействует с мутацией Ubx, вместе с которой они вызывают более выраженную трансформацию, чем каждая из них по отдельности. Гомозиготные особи Ubl/Ubl гибнут; выделены также другие летальные аллели Ubl, которые не вызывают никаких гомеозисных превращений и не взаимодействуют с мутацией Ubx. Гринлиф (Greenleaf) и его сотрудники провели биохимические исследования гена Ubl и показали, что аллель Ubl+ кодирует одну из субъединиц РНК-полимеразы II, необходимой для транскрипции генов, определяющих синтез матричных РНК (мРНК). Поскольку гомеозисным эффектом обладает лишь один аллель гена Ubl, а нуль-аллели им не обладают, можно заключить, что этот единственный гомеозисный аллель обусловливает синтез какого-то измененного фермента, который в свою очередь оказывает влияние на транскрипцию гена Ubx. Заключение о таком опосредованном эффекте недавно подтвердил Мортен, показав, что аллель Ubl оказывает действие на несколько других негомеозисных локусов по всему геному: просто локус Ubx обладает наибольшей чувствительностью к этому изменению фермента.
Все предшествующее обсуждение показывает, что необходимо провести исчерпывающий анализ генетических и морфогенетических свойств каждого локуса, прежде чем делать какие-либо общие выводы о механизме гомеозисных превращений, а следовательно, и о роли каждого отдельного гена в нормальном развитии. Такого рода обширное исследование было проведено на двух группах локусов; оно привело к заключению, что обе группы оказывают свое действие, участвуя в первоначальных детерминирующих событиях, контролирующих индивидуальность сегментов, т. е. специфическую дифференцировку каждого из них. Прежде чем обсуждать возможные механизмы, с помощью которых эти локусы вызывают трансформации, необходимо рассмотреть нормальный процесс сегментации у дрозофилы.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Спонтанные мутации
Спонтанные мутации О внезапных изменениях наследственности знал еще Дарвин. Взгляните на рисунок, где изображены овцы. Слева овца нормальная, в центре и справа овца и баран с сильно укороченными ногами. Сперва появилось одно-единственное такое животное. Изменение
Глава 5 СОМАТИЧЕСКИЕ МУТАЦИИ
Глава 5 СОМАТИЧЕСКИЕ МУТАЦИИ В гл. 4 мы высказали предположение, что соматические мутации V-генов играли важную роль в эволюции иммунной системы позвоночных. Сформулируем теперь несколько вопросов и постараемся ответить на них. Вопрос первый: каким образом создается
Микробы и мутации
Микробы и мутации Рассказ о трансформации, трансдукции и конъюгации должен был убедить читателя, что микроорганизмы действительно могут изменять свою природу. Во всех рассмотренных случаях микробиологи использовали влияние генетического материала (ДНК) одних микробов
ВТМ помогает изучать мутации
ВТМ помогает изучать мутации Вирус табачной мозаики, который уже не раз помогал нам разгадать многие загадки жизни, предлагается и в качестве модели для изучения мутаций. ВТМ содержит рибонуклеиновую кислоту, которая, как мы увидим далее, выполняет ту самую генетическую
Мутации вирусов в лаборатории
Мутации вирусов в лаборатории Мы говорили о мутантах ВТМ, созданных природой. Сегодня нам известны уже и его лабораторные мутанты. Первые из них увидели свет в лаборатории Г. Шрамма.Шрамм в своих опытах исходил из возможности замены аминогруппы (—NH2) в молекуле цитозина
Хромосомные аномалии и мутации
Хромосомные аномалии и мутации Факты, которые мы приводили выше, известны ученым уже довольно давно. Сопоставлением и изучением этих фактов и занимается генетика — наука о явлениях наследственности и изменчивости. Основное положение менделевской генетики — учение о
Полезные мутации переключателей
Полезные мутации переключателей Устойчивость к ядам, вирусам, бактериям и прочим паразитам, защитная окраска, превращение опадающих семян в неопадающие — все это примеры простых адаптаций, для развития которых бывает достаточно одной-двух удачных мутаций, поддержанных
5.1. Мутации
5.1. Мутации Теория мутаций составляет одну из основ генетики. Ее основные положения были разработаны голландским ученым Г. де Фризом еще в начале XX в.Мутации – это наследственные изменения генетического материала. Они характеризуются как редкие, случайные,
Мутации
Мутации Мутация — внезапное изменение гена. Она проявляется в первом же поколении потомков, если мутантный ген будет доминантным. Но рецессивный ген — мутант может скрытно наследоваться в течение нескольких поколений до тех пор, пока в родительскую пару не подберутся
Мутации и изменения, происходящие в онтогенезе
Мутации и изменения, происходящие в онтогенезе Если морфология представляет собой проявление сложного комплекса процессов развития, то эти процессы в свою очередь представляют собой проявление действия целого созвездия генов. Таково важное допущение, положенное в
Мутации с материнским эффектом
Мутации с материнским эффектом У таких разных организмов, как морские ежи и лягушки, события, происходящие на ранних стадиях дробления, и, в сущности, большая часть, если не все развитие, предшествующее гаструляции, не зависят от генома зиготы. Информацию, необходимую для
Мутации, затрагивающие органогенез
Мутации, затрагивающие органогенез Как мы убедились выше, события, происходящие на ранних стадиях развития, в значительной степени зависят от информации, поставляемой материнским организмом. Однако примерно ко времени гаструляции важную роль в дальнейшем развитии
Глава 8 Гомеозис в онтогенезе и филогенезе
Глава 8 Гомеозис в онтогенезе и филогенезе Смотри! Смотри! Что же я вижу? Лошадиную голову Там, где должен быть хвост! Руби Гольдберг
8.2. Мутации
8.2. Мутации Если бы организмы развивались поколение за поколением в одних и тех же окружающих условиях и передавали своему потомству все время одни и те же гены, сочетание эффектов генетической наследственности и морфического резонанса привело бы к бесконечному