5.1. Мутации
5.1. Мутации
Теория мутаций составляет одну из основ генетики. Ее основные положения были разработаны голландским ученым Г. де Фризом еще в начале XX в.
Мутации – это наследственные изменения генетического материала. Они характеризуются как редкие, случайные, ненаправленные события. Большая часть мутаций приводит к различным нарушениям нормального развития, некоторые из них летальны. Однако вместе с тем именно мутации являются исходным материалом для естественного отбора и биологической эволюции.
Частота мутаций возрастает под действием определенных факторов – мутагенов, способных изменять материал наследственности. В зависимости от их природы мутагены делятся на физические (ионизирующее излучение, УФ-излучение и др.), химические (большое число различных соединений), биологические (вирусы, мобильные генетические элементы, некоторые ферменты). Весьма условно деление мутагенов на эндогенные и экзогенные. Так, ионизирующее излучение, помимо первичного повреждения ДНК, образует в клетке нестабильные ионы (свободные радикалы), способные вторично вызывать повреждения генетического материала. Многие физические и химические мутагены являются также канцерогенами, т. е. индуцируют злокачественный рост клеток.
Частота мутаций подчиняется распределению Пуассона, применяемому в биометрии, когда вероятность отдельного события очень мала, а выборка, в которой может возникнуть событие, велика. Вероятность мутаций в отдельном гене довольно низкая, однако число генов в организме велико, а в генофонде популяции – огромно.
В литературе можно встретить различные классификации мутаций: по проявлению в гетерозиготе (доминантные, рецессивные), по инициирующему фактору (спонтанные, индуцированные), по локализации (генеративные, соматические), по фенотипическому проявлению (биохимические, морфологические, поведенческие, летальные и др.). На мой взгляд, эти показатели представляют собой скорее рабочие характеристики конкретных мутаций, а не основу для классификации.
Классифицируются мутации по характеру изменений генома. По этому показателю выделяют 4 группы мутаций, каждая из которых имеет многочисленные разновидности.
Генные мутации представляют собой изменения нуклеотидного состава ДНК отдельных генов. Мутационные изменения генов могут происходить в одной точке (односайтовые мутации) либо в нескольких разных точках (многосайтовые мутации). Термин «сайт» в генетике подразумевает определенное место в цепи молекулы ДНК. Современные методы молекулярной генетики позволили определить два основных процесса формирования генных мутаций: замена нуклеотидов и сдвиг рамки считывания, каждый из которых имеет свои варианты (рис. 5.1).
Транзиции – при замене сохраняется месторасположение пуриновых и пиримидиновых нуклеотидов (АТ ? ГЦ, ГЦ ? АТ и т. п.).
Трансверсии – при замене пуриновые и пиримидиновые нуклеотиды меняются местами (АТ ? ЦГ, АТ ? ТА и т. п.).
Мутации вследствие сдвига рамки считывания встречаются более часто. Они проявляются в двух вариантах: инсерция (вставка) и делеция (утеря) одного или нескольких нуклеотидов. Необходимо отметить, что вставка сдвигает рамку считывания в одном направлении, а делеция – в противоположном.
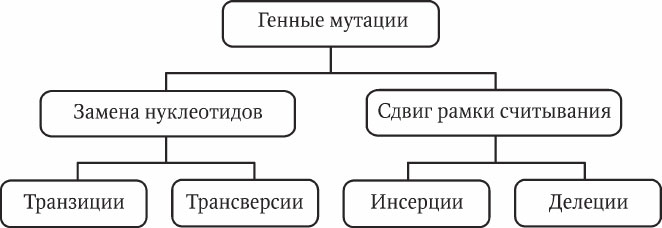
Рис. 5.1. Генные мутации
Механизм возникновения генных мутаций наиболее детально изучен у вирусов и бактерий. Согласно концепции американского генетика Р. фон Борстела, генные мутации возникают в результате ошибок «трех Р»: репликации, репарации и рекомбинации.
В процессе репликации возможна замена нуклеотидов вследствие некоторой неоднозначности принципа комплементарности. Азотистые основания нуклеотидов ДНК могут существовать в нескольких таутомерных формах. Таутомеризация – это изменение положения водорода в молекуле, меняющее ее химические свойства. Некоторые таутомеры нуклеотидов меняют способность формировать водородные связи с другими нуклеотидами. У аналогов нуклеотидов таутомерия происходит значительно чаще, чем у типичных форм, что объясняет их мутагенный эффект. Примером может служить аналог тимидина 5-бромурацил, который способен в некоторых таутомерных формах вступать в связь с гуанином. Вследствие вырожденности генетического кода замены нуклеотидов могут иметь различные фенотипические проявления. Они могут не вызвать замены аминокислот или привести к появлению аминокислоты, близкой по своим свойствам, но могут привести к замене на аминокислоту с другими физико-химическими свойствами или к образованию стоп-кодона.
Большинство мутаций со сдвигом рамки считывания обнаружены в участках ДНК, состоящих из одинаковых нуклеотидов. Существует гипотеза возникновения этих мутаций вследствие диссоциации и неправильного восстановления нитей в данных участках.
Резкий рост мутаций при нарушении системы репарации и взаимосвязь мутационного и рекомбинационного процессов продемонстрированы в многочисленных исследованиях. Процессы репарации и рекомбинации мы рассмотрим ниже.
Хромосомные мутации (аберрации) – это изменения структуры хромосом: внутрихромосомные или межхромосомные перестройки, возникающие при разрывах хромосом. Хромосомные перестройки обычно приводят к различным фенотипическим проявлениям. Выделяют следующие виды аберраций (рис. 5.2).
Дупликация – дублирование участка хромосомы.
Амплификация – многократное повторение участка хромосомы.
Повторы генетического материала не оказывают такого отрицательного влияния на организм как делеции и дефишенси. Показана значительная роль дупликаций в эволюции генома, поскольку они создают дополнительные участки генетического материала, доступные для мутирования, изменения функций генов и естественного отбора.
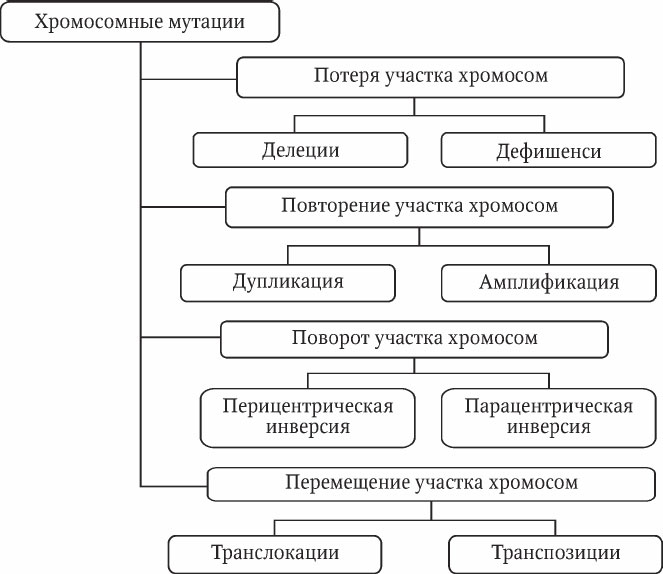
Рис. 5.2. Хромосомные мутации
Явление амплификации можно наблюдать при культивировании клеток с различными повреждающими агентами, но оно встречается и в природе как закономерный процесс онтогенеза, когда необходимо резко увеличить экспрессию каких-либо генов. В последнем случае возможны два варианта: либо амплифицированная ДНК остается связанной с хромосомой, образуя многочисленные репликативные вилки (например, в фолликулярных клетках дрозофилы); либо синтезированная ДНК отделяется от материнской и многократно реплицируется (как ДНК, содержащая геныр-РНК ооцитов амфибий).
Инверсия – поворот участка хромосомына 180°. Инверсия приводит к изменению линейной последовательности генов. Она встречается в двух вариантах: перицентрическая инверсия (центромера входит в инвертированный участок) и парацентрическая инверсия (центромера не входит в инвертируемый участок). Негативный эффект инверсии зависит от локализации точек разрывов, их близости к жизненно важным генам. Необходимо отметить, что инверсии встречаются в природных популяциях чаще других хромосомных перестроек. Они представляют собой распространенный путь преобразований генетического материала в эволюции, являясь факторами изоляции и дивергенции новых форм в пределах вида. Реципрокные транслокации – обмен участками хромосом между негомологичными хромосомами. В результате такой транслокации изменяется характер сцепления генов – гены, принадлежащие к разным хромосомам, могут наследоваться как одна группа сцепления. Характер конъюгации при транслокации меняется – вместо бивалентов образуется квадривалент в виде «фигуры креста» (рис. 5.3).
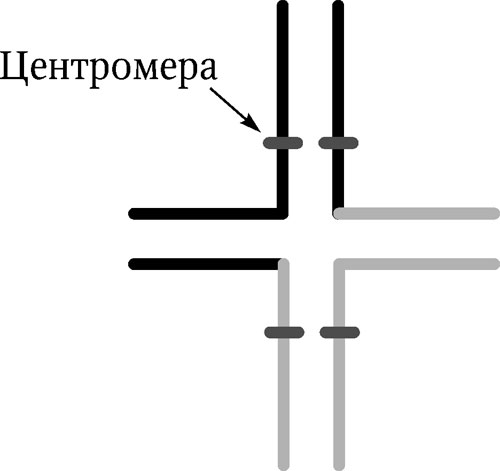
Рис. 5.3. Синапсис хромосом при реципрокной транслокации в профазе мейоза. На каждой хромосоме отмечена центромера
Гетерозиготы по реципрокным транслокациям обладают пониженной плодовитостью, так как продуцируют дефектные гаметы. Только у двух видов гамет из шести возможных при разных способах расхождения хромосом имеются полные комплекты генов. Остальные несут дупликации и нехватки и не могут дать жизнеспособного потомства. У животных реципрокные транслокации встречаются редко, обычно с негативным эффектом, но они широко распространены у растений. Благодаря специальным механизмам, транслокация обеспечивает изоляцию новых форм.
Участок хромосомы может менять свое положение и без реципрокного (взаимного) обмена, оставаясь на той же хромосоме или включаясь в другую. Такое явление называется транспозицией. Транспозиции будут рассматриваться ниже как важный самостоятельный раздел современной генетики.
Вероятно, все типы хромосомных перестроек имеют единый механизм и обусловлены лабильностью генома.
Причиной изменения фенотипа при различных хромосомных перестройках часто является изменение расположения гена. Этот феномен получил название эффект положения гена. Он показан для многих генов и обычно влияет на их регуляторную систему. Например, при перемещении гена из эухроматина в гетерохроматиновую область его активность утрачивается, хотя сам ген не изменяется.
Геномные мутации – это изменения числа хромосом. Среди геномных мутаций также выделяют несколько разновидностей (рис. 5.4).
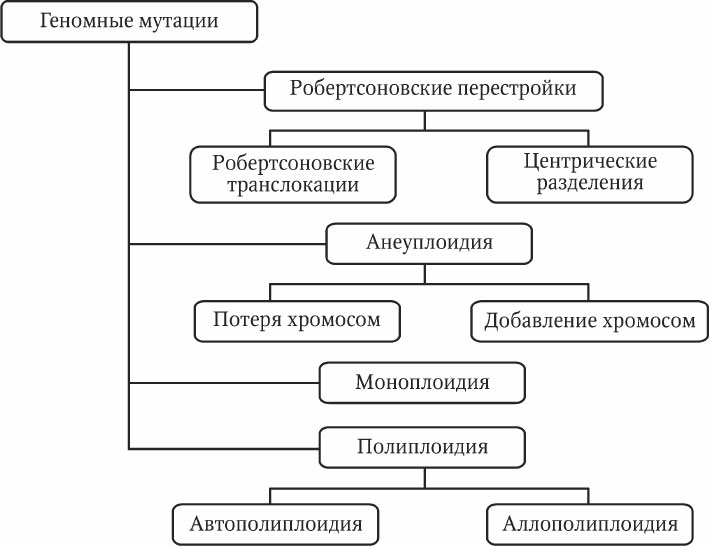
Рис. 5.4. Геномные мутации
Робертсоновские перестройки – слияния и разделения хромосом в области центромеры. Названы они по имени В. Робертсона, который предложил свою гипотезу механизма таких мутаций. Центрические слияния (робертсоновские транслокации) представляют собой слияния двух негомологичных акроцентрических хромосом с образованием одной субметацентрической хромосомы. При разделении, наоборот, одна субметацентрическая хромосома делится на две акроцентрические хромосомы. При этом должна образоваться новая центромера, иначе хромосома без центромеры будет потеряна при митозе.
Робертсоновские перестройки приводят к изменению числа хромосом в кариотипе, не влияя на общее количество генетического материала в клетке. Оба варианта перестроек представлены в природе, но робертсоновские транслокации встречаются значительно чаще. Они являются одним из магистральных путей эволюции кариотипов.
Анеуплоидия – изменение числа хромосом, не кратное гаплоидному набору. Как правило, представляет собой добавление или потерю одной или двух хромосом диплоидного набора. У животных анеуплоидия обычно приводит к тяжелым аномалиям или летальности. Однако у растений трисомия (наличие трех гомологичных хромосом) может служить фактором генетического разнообразия. Причиной анеуплоидии является нерасхождение хромосом в мейозе и образование несбалансированных гамет.
Моноплоидия (гаплоидия) – гаплоидное состояние диплоидного организма. Эта мутация интенсивно изучается у растений, так как позволяет видеть проявление рецессивных аллелей. У животных моноплоидия обычно приводит к летальному исходу.
Автополиплоидия – наличие в клетке более двух одинаковых гаплоидных наборов. Эта разновидность мутации довольно широко представлена в природе у протистов, грибов и растений. Плоидность макронуклеуса инфузорий может достигать нескольких сотен. У животных встречается редко и обычно приводит к летальному исходу на ранних стадиях эмбриогенеза. У культурных растений сбалансированные полиплоиды (т. е. кариотипы с четным числом гаплоидных наборов – 4n, 6n, 8n и т. п.) получают искусственным путем из-за их более крупных размеров. Несбалансированные полиплоиды (3n, 5n, 7n и т. п.) растений часто имеют пониженную фертильность вследствие нарушений мейоза. Тем не менее некоторые растения-триплоиды (3n) обладают большими размерами и продуктивностью по сравнению с диплоидными (2n) и тетраплоидными (4n).
Аллополиплоидия – объединение в клетке разных геномов посредством гибридизации. В природе для многих цветковых растений описаны полиплоидные ряды различной степени плоидности. Эти ряды возникают путем гибридизации разных видов и последующего удвоения родительских гаплоидных наборов. Так преодолевается барьер бесплодия при скрещивании разных видов.
Цитоплазматические мутации – это изменения неядерных генов. Генетическая информация органелл (в отличие от хромосомной) представлена в зиготе в разном объеме от каждого из родителей. Чаще всего преобладает вклад материнского организма, но у некоторых видов наследуются внеядерные геномы от обоих родителей. Мутации генома органелл обычно представлены точечными мутациями и делециями. Основными носителями внеядерной генетической информации в клетках высших эукариот являются хлоропласты и митохондрии.
В хлоропластах фенотипическое проявление мутаций выражается нарушением фотосинтеза, изменением чувствительности к температуре и устойчивости к антибиотикам. Генетика хлоропластов разработана хуже, чем генетика митохондрий.
У позвоночных митохондрии служат единственным хранилищем внеядерного генетического материала. Источником митохондрий в зиготе, вероятно, служит только яйцеклетка. Хотя 95 % митохондриальных белков кодируется в ядре, мутации мт-ДНК происходят в 10 раз чаще, чем в ядерных генах, поэтому их вклад ощутим.
Фенотипическое проявление мутаций мт-ДНК во многом зависит от уровня потребления АТФ той или иной тканью. Поскольку наиболее энергозависимыми являются нервная и мышечная системы, такие мутации наиболее часто проявляются в форме различных нейропатий и миопатий.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Спонтанные мутации
Спонтанные мутации О внезапных изменениях наследственности знал еще Дарвин. Взгляните на рисунок, где изображены овцы. Слева овца нормальная, в центре и справа овца и баран с сильно укороченными ногами. Сперва появилось одно-единственное такое животное. Изменение
Глава 5 СОМАТИЧЕСКИЕ МУТАЦИИ
Глава 5 СОМАТИЧЕСКИЕ МУТАЦИИ В гл. 4 мы высказали предположение, что соматические мутации V-генов играли важную роль в эволюции иммунной системы позвоночных. Сформулируем теперь несколько вопросов и постараемся ответить на них. Вопрос первый: каким образом создается
Микробы и мутации
Микробы и мутации Рассказ о трансформации, трансдукции и конъюгации должен был убедить читателя, что микроорганизмы действительно могут изменять свою природу. Во всех рассмотренных случаях микробиологи использовали влияние генетического материала (ДНК) одних микробов
ВТМ помогает изучать мутации
ВТМ помогает изучать мутации Вирус табачной мозаики, который уже не раз помогал нам разгадать многие загадки жизни, предлагается и в качестве модели для изучения мутаций. ВТМ содержит рибонуклеиновую кислоту, которая, как мы увидим далее, выполняет ту самую генетическую
Мутации вирусов в лаборатории
Мутации вирусов в лаборатории Мы говорили о мутантах ВТМ, созданных природой. Сегодня нам известны уже и его лабораторные мутанты. Первые из них увидели свет в лаборатории Г. Шрамма.Шрамм в своих опытах исходил из возможности замены аминогруппы (—NH2) в молекуле цитозина
Хромосомные аномалии и мутации
Хромосомные аномалии и мутации Факты, которые мы приводили выше, известны ученым уже довольно давно. Сопоставлением и изучением этих фактов и занимается генетика — наука о явлениях наследственности и изменчивости. Основное положение менделевской генетики — учение о
Полезные мутации переключателей
Полезные мутации переключателей Устойчивость к ядам, вирусам, бактериям и прочим паразитам, защитная окраска, превращение опадающих семян в неопадающие — все это примеры простых адаптаций, для развития которых бывает достаточно одной-двух удачных мутаций, поддержанных
5.1. Мутации
5.1. Мутации Теория мутаций составляет одну из основ генетики. Ее основные положения были разработаны голландским ученым Г. де Фризом еще в начале XX в.Мутации – это наследственные изменения генетического материала. Они характеризуются как редкие, случайные,
Мутации
Мутации Мутация — внезапное изменение гена. Она проявляется в первом же поколении потомков, если мутантный ген будет доминантным. Но рецессивный ген — мутант может скрытно наследоваться в течение нескольких поколений до тех пор, пока в родительскую пару не подберутся
Мутации и изменения, происходящие в онтогенезе
Мутации и изменения, происходящие в онтогенезе Если морфология представляет собой проявление сложного комплекса процессов развития, то эти процессы в свою очередь представляют собой проявление действия целого созвездия генов. Таково важное допущение, положенное в
Мутации с материнским эффектом
Мутации с материнским эффектом У таких разных организмов, как морские ежи и лягушки, события, происходящие на ранних стадиях дробления, и, в сущности, большая часть, если не все развитие, предшествующее гаструляции, не зависят от генома зиготы. Информацию, необходимую для
Мутации, затрагивающие органогенез
Мутации, затрагивающие органогенез Как мы убедились выше, события, происходящие на ранних стадиях развития, в значительной степени зависят от информации, поставляемой материнским организмом. Однако примерно ко времени гаструляции важную роль в дальнейшем развитии
Мутации, воздействующие на развитие определенных органов
Мутации, воздействующие на развитие определенных органов В то время как локус Т оказывает, по-видимому, самые разнообразные воздействия на все развитие эктодермы, существуют мутации, вызывающие более специфические дефекты. Примером служит мутация cardiac lethal (с) у аксолотля
Гомеозис и гомеозисные мутации
Гомеозис и гомеозисные мутации Действие генов теснейшим образом связано с онтогенезом, и эта их связь выявляется при возникновении мутаций, которые резко прерывают развитие организма. Существуют, однако, мутации другого класса, которые изменяют процесс онтогенеза, но