Полезные мутации переключателей
Полезные мутации переключателей
Устойчивость к ядам, вирусам, бактериям и прочим паразитам, защитная окраска, превращение опадающих семян в неопадающие — все это примеры простых адаптаций, для развития которых бывает достаточно одной-двух удачных мутаций, поддержанных отбором. Более глубокие преобразования складываются из десятков и сотен подобных «мелочей». Одна простая полезная мутация может изменить ландшафт приспособленности для организма (или вывести его в новую область этого ландшафта) — например, изменить взаимоотношения организма со средой, сделав возможной жизнь в условиях, доселе неприемлемых, — и в результате какие-то другие мутации, прежде бывшие вредными, станут полезными и рано или поздно зафиксируются, открыв возможности для новых изменений.
Поскольку для крупных эволюционных преобразований требуется последовательное закрепление множества мутаций, такие изменения трудно «расшифровать» на молекулярном уровне, а уж пронаблюдать воочию и вовсе невозможно из-за длительности процесса. Те случаи, которые все-таки удается расшифровать, — это, по-видимому, редкие, нетипичные случаи, когда крупное изменение обеспечивается всего несколькими мутациями. Но такие случаи есть, и мы пока не знаем наверняка, так ли уж мала их роль на больших — эволюционно значимых — отрезках времени. Мы познакомимся с одним из таких случаев, когда всего две мутации обеспечили интересное и важное адаптивное приобретение у многоклеточного животного — нематоды (круглого червя). В результате двух мутаций нематоды приобрели способность к самооплодотворению — стали гермафродитами. Этот пример важен нам также для того, чтобы показать, как в ходе эволюции подстраиваются друг к другу разные гены, регулирующие определенную функцию.
У большинства нематод, как и у многих других животных, пол определяется генетически, при помощи половых хромосом. Если в оплодотворенном яйце две X-хромосомы, из яйца разовьется самка, если одна — самец (Haag, 2005). Однако у двух наиболее изученных видов нематод, Caenorhabditis elegans и C. briggsae, особи с двумя X-хромосомами — не самки, а гермафродиты. Их гонады (половые железы) на поздних личиночных стадиях производят спермии. Нематоды утратили в ходе эволюции жгутики, поэтому и спермии у них безжгутиковые. Они похожи на амеб и передвигаются при помощи псевдоподий. У гермафродитов безжгутиковые спермии поздних личинок переползают в специальные хранилища — сперматеки — и здесь ждут своего часа. Гонады взрослых гермафродитов производят уже не спермии, а яйцеклетки. Они могут быть оплодотворены как собственными спермиями из сперматеки, так и спермиями самца в результате спаривания.
Предки C. elegans и C. briggsae были нормальными раздельнополыми червями. Это следует, в частности, из того, что все прочие виды рода Caenorhabditis — раздельнополые. По-видимому, гермафродитизм у C. elegans и C. briggsae является «эволюционно молодым», новым признаком. Чтобы разобраться, как он возник, понадобились выдумка, знание геномов нескольких видов нематод и эксперименты, осуществленные американскими биологами (Baldi et al., 2009). Вот как это было.
Для начала уточним, какие гены работают при формировании половых различий у нематод. Наличие одной (а не двух) X-хромосом у самцов служит триггером, включающим синтез белка HER-1, который ингибирует белок TRA-2. Подавление активности TRA-2 через пару промежуточных шагов приводит к уничтожению белка TRA-1, функция которого состоит в том, чтобы отключать ряд ключевых генов, направляющих развитие по «мужскому» пути (в том числе ген fog-3, контролирующий сперматогенез)[29]. У гермафродитов белок HER-1 не образуется, но они все равно производят спермии, потому что активность гена tra-2 на стадии личинки подавляется белками FOG-2 и GLD-1. Таким образом, гермафродиты получаются из-за введения в игру дополнительного правила, связанного с регуляцией гена tra-2.
Запутались? Ничего страшного, это в порядке вещей. Регуляторные сети, управляющие развитием животных, как правило, сложны, громоздки и неоптимальны[30]. К счастью, их не нужно учить наизусть никому, кроме студентов соответствующих специальностей. Чтобы облегчить дело, можно представить устрашающее описание из предыдущего абзаца в виде графической схемы (см. следующую страницу).
На таких схемах стрелочками обозначают положительные влияния (включение гена, активация белка), а «тупичками» — отрицательные (ингибирование белка, подавление активности гена).
Так вот, зная все это, исследователи подумали, что можно попробовать превратить самок раздельнополого вида C. remanei в гермафродитов, подавив активность какого-нибудь гена, блокирующего сперматогенез. Например, гена tra-2.
Начали они с того, что отключили tra-2 при помощи РНК-интерференции[31]. В результате из яиц с двумя X-хромосомами развились вместо самок обыкновенные самцы, производящие нормальные спермии, но никаких яйцеклеток.
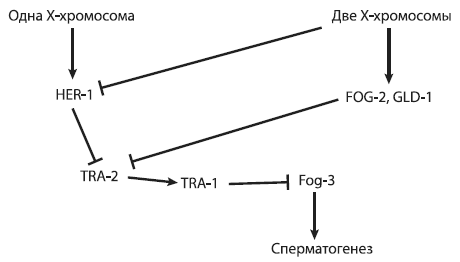
Схема регуляции развития пола у нематоды C. elegans, у которой вместо самок гермафродиты.
Тогда ученые справедливо рассудили, что они, видимо, перестарались. Ведь у настоящих гермафродитов активность гена tra-2 хоть и снижена, но не до нуля. Когда эту ситуацию удалось воспроизвести у C. remanei, уменьшив экспрессию tra-2, на свет появились животные, которых авторы назвали псевдогермафродитами. Тело у них было «женское», но в гонадах формировались одновременно и яйцеклетки, и спермии. Правда, эти существа оказались неспособны к самооплодотворению (отсюда и приставка «псевдо»). После скрещивания с самцом превдогермафродиты откладывали оплодотворенные яйца, хоть и в меньшем количестве, чем обычные самки. Это значит, что яйцеклетки у них более или менее нормальные, а неспособность к самооплодотворению, скорее всего, объясняется дефектами спермиев.
Действительно, спермии псевдогермафродитов оказались нормальны по всем параметрам, кроме одного: они неактивны, никуда не ползут, и в частности не перебираются в сперматеки.
У обычных нематод спермии активируются (начинают ползать) под воздействием белков, содержащихся в семенной жидкости. Сохранили ли спермии псевдогермафродитов способность активироваться под действием этих белков? Чтобы это проверить, авторы скрещивали псевдогермафродитов с самцами C. elegans. Самцы C. elegans охотно спариваются с самками C. remanei, но эти браки бесплодны (как-никак два вида разошлись, судя по показаниям молекулярных часов, около 80 млн лет назад, а для развития генетической несовместимости обычно хватает нескольких миллионов лет[32]). Идея такого спаривания как раз и состояла в том, чтобы половые клетки от разных видов не сливались, но при этом псевдогермафродиты могли получить нормальные белки семенной жидкости.
И когда после этого адюльтера псевдогермафродиты C. remanei отложили оплодотворенные яйца, некоторые из них оказались жизнеспособными! Из них вывелись нормальные самки C. remanei (не псевдогермафродиты, потому что им никто не подавлял активность гена tra-2). Это означает, что спермии у псевдогермафродитов получились нормальными, не хватает только активирующих белков.
Это открытие сузило круг потенциальных генов-мишеней, на которые нужно воздействовать, чтобы превратить псевдогермафродитов в полноценных гермафродитов. «Подходящим» геном оказался swm-1, который кодирует белок, подавляющий активность других белков — протеаз, активирующих спермии. Ген swm-1 отвечает за предотвращение преждевременной активации спермиев у самцов C. elegans, но у него есть и другие функции. Ученые понизили активность этого гена у псевдогермафродитов — и те, к большой радости исследователей, приобрели способность к самооплодотворению.
Таким образом, для появления нового признака — гермафродитизма — достаточно уменьшить активность двух генов, входящих в два разных регуляторных каскада. Для этого нужны две мутации. Изменение активности того или иного гена в результате случайной мутации — дело самое обычное. Проблема в другом: одновременное возникновение двух мутаций, полезных только вместе, но не по отдельности, крайне маловероятно. Могли ли они появиться последовательно или это равносильно преодолению пропасти в два прыжка? Исследователи рассмотрели два возможных сценария.
1. Сначала произошла мутация, уменьшившая активность swm-1 у самок, что позволяет активировать собственные спермии (если они есть). Эта мутация поначалу была нейтральной, потому что самки еще не умели производить спермии. В дальнейшем возникла мутация, понизившая активность tra-2. Эта мутация сразу оказалась полезной (т. е. была поддержана отбором), потому что превратила самок в самодостаточных гермафродитов (о полезности гермафродитизма см. ниже). Но могла ли первая мутация не принести вреда, если известно, что ген swm-1 многофункционален? И вот тут-то на помощь приходит знание геномов изучаемых червей (недаром в последние годы биологи столько сил и средств тратят на прочтение геномов всевозможных тварей). Оказывается, в геномах раздельнополых видов есть только одна копия swm-1, а у гермафродитных есть еще и вторая, немного отличающаяся копия. По-видимому, становлению гермафродитизма способствовала дупликация (удвоение) этого гена, что и позволило снять «адаптивный конфликт». Одна из копий изменилась, чтобы обеспечить активацию спермиев у гермафродитов, а вторая продолжила выполнение остальных функций[33].
2. Второй сценарий начинается с мутации, снизившей активность tra-2. Это привело к появлению псевдогермафродитов. Лишь после этого возникла и закрепилась мутация, снижающая активность swm-1. Однако псевдогермафродиты фактически являются всего лишь неполноценными самками: к самооплодотворению они не способны, а с ролью самок справляются хуже, чем настоящие самки. Поэтому на первый взгляд кажется, что отбор должен был отсеять первую мутацию. Но вспомним, что самки C. remanei привлекательны для самцов других видов, а спаривание с этими самцами дает им возможность самооплодотвориться. Эта особенность могла стать «мостиком», с помощью которого эволюционирующий вид сумел преодолеть опасный промежуточный этап и дождаться возникновения второй мутации. В некоторых ситуациях псевдогермафродиты, способные к самооплодотворению после спаривания с самцами других видов, могли иметь адаптивное преимущество — например, когда численность популяции критически снижалась.
Таким образом, при ближайшем рассмотрении оказывается, что червям не нужно было «преодолевать пропасть в два прыжка». Там все-таки были мостики.
Изменение активности гена в сторону уменьшения или увеличения может произойти в результате самых разных мутаций. Это могли быть мутации в регуляторных областях самих генов tra-2 и swm-1, или мутации генов-регуляторов, управляющих их работой, или мутации регуляторов регуляторов, и т. д. Важно, что в такой ситуации — когда оказывается выгодно уменьшить или увеличить экспрессию какого-либо гена — вероятность того, что случайная мутация, меняющая активность гена, окажется полезной, а не вредной, приближается к 1/2, т. е. становится чрезвычайно высокой. Скорее всего, именно поэтому очень многие «прогрессивные» эволюционные преобразования, как теперь выясняется, были связаны с изменениями уровня активности генов, а не их белок-кодирующих последовательностей. Между прочим, это относится и к эволюции человека (Gilad et al., 2006).
Появление новых признаков путем изменения активности генов — один из магистральных путей эволюции. Почему? Да прежде всего потому, что это просто. В инструкциях по устранению неисправностей в работе электроприборов неизменно присутствует «мудрый совет», раздражающий многих: проверьте, включена ли вилка в розетку. За ним кроется универсальный принцип: во многих случаях нужного эффекта можно добиться, манипулируя только выключателями и не развинчивая весь механизм.
Конкретные мутации, ответственные за снижение активности генов tra-2 и swm-1 у предков C. elegans и C. briggsae, возможно, были разными. Например, известно, что у гермафродитов первого вида в подавлении активности tra-2 участвует ген fog-2, отсутствующий у второго вида. Предки C. briggsae утратили ген fog-2, вероятно за ненадобностью. У этого вида ключевую роль в обеспечении нормального сперматогенеза у гермафродитов играет другой ген — she-1. Этот пример показывает, что в ходе эволюции «переключатели» генетических регуляторных каскадов могут меняться, в то время как структура и функции каскадов остаются прежними.
Это относится и к механизмам определения пола. Разделение на самцов и самок есть у большинства животных. Соответственно, есть и генетические «программы» (большие и сложные) развития по мужскому и женскому пути. Однако переключатели, направляющие развитие по одному из двух путей, часто меняются в ходе эволюции. Например, у тех же нематод многие виды перешли от хромосомной детерминации пола к «средовой», т. е. пол у них зависит не от генов, а от условий, в которых проходит развитие. Мы теперь понимаем, что такие эволюционные изменения относятся к числу высоковероятных. Точно так же вы можете заменить кнопку электрического выключателя в своей комнате на систему последовательно и параллельно соединенных тумблеров, реле или повесить шнурок с кисточкой, не меняя при этом проводку и люстру. Управлять освещенностью, возможно, станет удобнее, но сам «фенотип» от этого не изменится: свет по-прежнему будет или включен, или выключен.
Ну и последнее: зачем вообще кому-то понадобилось переходить к гермафродитизму? Как могли мутации, превратившие самок в гермафродитов, оказаться полезными для предков C. elegans и C. briggsae? Дело в том, что в некоторых ситуациях гермафродитизм дает очевидное преимущество[34]. Например, если участки, пригодные для жизни нематод данного вида, встречаются очень редко (это вполне справедливо для представителей рода Caenorhabditis) и вероятность того, что хотя бы один червь попадет на новый, незаселенный участок, очень мала, то вероятность того, что на этот участок попадут сразу два червя — самец и самка, — будет и вовсе ничтожной. Естественно, в такой ситуации преимущество получат черви, способные размножаться без посторонней помощи, путем самооплодотворения. Или, на худой конец, с помощью самцов других видов.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
Спонтанные мутации
Спонтанные мутации О внезапных изменениях наследственности знал еще Дарвин. Взгляните на рисунок, где изображены овцы. Слева овца нормальная, в центре и справа овца и баран с сильно укороченными ногами. Сперва появилось одно-единственное такое животное. Изменение
Глава 5 СОМАТИЧЕСКИЕ МУТАЦИИ
Глава 5 СОМАТИЧЕСКИЕ МУТАЦИИ В гл. 4 мы высказали предположение, что соматические мутации V-генов играли важную роль в эволюции иммунной системы позвоночных. Сформулируем теперь несколько вопросов и постараемся ответить на них. Вопрос первый: каким образом создается
Микробы и мутации
Микробы и мутации Рассказ о трансформации, трансдукции и конъюгации должен был убедить читателя, что микроорганизмы действительно могут изменять свою природу. Во всех рассмотренных случаях микробиологи использовали влияние генетического материала (ДНК) одних микробов
ВТМ помогает изучать мутации
ВТМ помогает изучать мутации Вирус табачной мозаики, который уже не раз помогал нам разгадать многие загадки жизни, предлагается и в качестве модели для изучения мутаций. ВТМ содержит рибонуклеиновую кислоту, которая, как мы увидим далее, выполняет ту самую генетическую
Мутации вирусов в лаборатории
Мутации вирусов в лаборатории Мы говорили о мутантах ВТМ, созданных природой. Сегодня нам известны уже и его лабораторные мутанты. Первые из них увидели свет в лаборатории Г. Шрамма.Шрамм в своих опытах исходил из возможности замены аминогруппы (—NH2) в молекуле цитозина
1. Полезные ранения
1. Полезные ранения Когда приходится знакомиться с теми представлениями о жизни природы, которые имели наши предки две-три тысячи лет тому назад, поражает смесь верной, иногда довольно тонкой наблюдательности с самыми странными предрассудками и фантастическими
1. Полезные ранения
1. Полезные ранения Когда приходится знакомиться с теми представлениями о жизни природы, которые имели наши предки две-три тысячи лет тому назад, поражает смесь верной, иногда довольно тонкой наблюдательности с самыми странными предрассудками и фантастическими
Глава 2 Полезные ошибки
Глава 2 Полезные ошибки Все здание теории эволюции, а значит и биологии в целом, основано на том, что некоторые случайные наследственные изменения оказываются полезными, т. е. повышают эффективность размножения (по-другому называемую приспособленностью). Термин
5.1. Мутации
5.1. Мутации Теория мутаций составляет одну из основ генетики. Ее основные положения были разработаны голландским ученым Г. де Фризом еще в начале XX в.Мутации – это наследственные изменения генетического материала. Они характеризуются как редкие, случайные,
Мутации
Мутации Мутация — внезапное изменение гена. Она проявляется в первом же поколении потомков, если мутантный ген будет доминантным. Но рецессивный ген — мутант может скрытно наследоваться в течение нескольких поколений до тех пор, пока в родительскую пару не подберутся
Мутации с материнским эффектом
Мутации с материнским эффектом У таких разных организмов, как морские ежи и лягушки, события, происходящие на ранних стадиях дробления, и, в сущности, большая часть, если не все развитие, предшествующее гаструляции, не зависят от генома зиготы. Информацию, необходимую для
Мутации, затрагивающие органогенез
Мутации, затрагивающие органогенез Как мы убедились выше, события, происходящие на ранних стадиях развития, в значительной степени зависят от информации, поставляемой материнским организмом. Однако примерно ко времени гаструляции важную роль в дальнейшем развитии
Гомеозис и гомеозисные мутации
Гомеозис и гомеозисные мутации Действие генов теснейшим образом связано с онтогенезом, и эта их связь выявляется при возникновении мутаций, которые резко прерывают развитие организма. Существуют, однако, мутации другого класса, которые изменяют процесс онтогенеза, но
Роль двоичных переключателей в интерпретации позиционной информации
Роль двоичных переключателей в интерпретации позиционной информации Возможный характер генетической реакции на позиционную информацию был описан в некотором формальном смысле Кауфманом (Kauffmann). Согласно его модели, гены, реагирующие на позиционную информацию,
8.2. Мутации
8.2. Мутации Если бы организмы развивались поколение за поколением в одних и тех же окружающих условиях и передавали своему потомству все время одни и те же гены, сочетание эффектов генетической наследственности и морфического резонанса привело бы к бесконечному