Глава 4 Эволюция и структурная организация яиц и зародышей
Глава 4
Эволюция и структурная организация яиц и зародышей
Много еще и теперь из нее (Земли) выходит животных,
Влагой дождей воплощенных и жаром горячего солнца.
Не мудрено, что крупней были твари тогда, да и больше
Их порождалось, землей молодой и эфиром взращенных...
Ибо в полях и тепла изобилие было и влаги.
Всюду поэтому, где предоставлялось удобное место,
В почву корнями вцепясь, вырастали утробы; когда же
Их разверзали, созрев в урочное время, младенцы...
Тит Лукреций Кар «О природе вещей»
Пространственная организация и начало морфогенеза
Сложные изменения размеров и формы, которые составляют морфологическую эволюцию, могут происходить довольно быстро и достигаются главным образом в результате изменений не структурных генов, а регуляторных элементов, определяющих процесс развития. В дальнейших главах мы еще вернемся к природе этих регуляторных элементов и их эволюции. Здесь же нам необходимо рассмотреть те процессы, с помощью которых информация, закодированная в геноме, экспрессируется во все возрастающей морфологической сложности развивающегося зародыша.
Яйцо - это не просто изотропный реакционный сосуд, содержащий ДНК и другие компоненты, необходимые для транскрипции и трансляции. Яйцо обладает внутренней структурой, которая так же непосредственно, как и ДНК, участвует в определении процессов эмбрионального морфогенеза. Сама инициация процесса развития и изменений во внутренней организации яйца в то время, когда оно начинает дробиться в ответ на активацию спермой, послужило источником для некоторых из самых глубоких разногласий в истории становления эмбриологии как науки.
Ранние эмбриологи столкнулись с кажущимся парадоксом эпигенеза - возникновением животного организма с его сложной структурой из яйца, кажущегося бесструктурным. Или же, как в 1764 г. сформулировал эту проблему ведущий теоретик эмбриологии XVIII в. Шарль Бонне: «Если организованные тела не преформированы, то в таком случае их формирование должно происходить ежедневно, подчиняясь законам какой-то особой механики. Так вот, скажите мне, пожалуйста, какая механика может управлять формированием головного мозга, сердца, легкого и многих других органов?» Существовало лишь одно логическое решение этой дилеммы. Бонне высказал мнение, что в яйце преформирован, заложен в готовом виде, миниатюрный, но вполне сформированный организм, который во время развития лишь увеличивается в размерах, но не становится сложнее. Нам такая идея кажется странной, да она и в самом деле повлекла за собой немало нелепостей, таких как теория вложения, согласно которой у зародыша, изначально содержащегося в яйце, уже есть яичник с яйцами, содержащими еще более мелкие зародыши, и т.д. Делались даже попытки вычислить, сколько зародышей было заключено в яичниках Евы.
Теория преформации не утверждала, что содержащийся в яйце крошечный зародыш-предшественник - это гомункулюс, идентичный по своей структуре той взрослой особи, в которую ему суждено превратиться; она лишь настаивала на том, что преформированные структуры присутствуют в яйце в виде некой организованной сущности, способной к развертыванию с выявлением взрослой формы. Однако идеи Бонне не смогли противостоять становлению эмбриологии как науки, о чем свидетельствует работа К. Бэра (von Baer), показавшего, что морфологическая сложность зародышей возрастает по мере их развития. К. Бэр продемонстрировал, что все дифференцированные ткани позвоночных возникают из трех морфологически простых зародышевых листков: центральная нервная система и покровные ткани - из эктодермы, мышцы и скелет - из мезодермы, а органы пищеварения - из энтодермы. Позднее, в XIX в., Ковалевский показал, что и сами зародышевые листки возникают эпигенетически.
На смену строгому преформизму пришла столь же строгая теория эпигенеза, согласно которой сложные структуры возникают de novo из бесструктурного яйца. Такое представление в свою очередь вскоре было опровергнуто. В конце XIX в. по мере увеличения числа и детальности исследований раннего развития зародышей различных морских животных становилось все яснее, что яйцо содержит определенную морфогенетическую информацию и что разные участки яйца неравноценны по своим потенциям к развитию. Это не был преформизм в понимании Бонне. Сторонники этого нового течения не утверждали, что в яйце заключен организм в миниатюре, но и не считали яйцо бесструктурным. Уже в 1877 г. Ланкестер (Е. Lankester) выдвинул гипотезу, сформулированную в молекулярных терминах:
«Любая дифференцировка клеток, развитие клеток одного типа из клеток другого типа, зависит от внутреннего перемещения физиологических молекул, составляющих протоплазму таких клеток... Эти молекулы... находятся в клетке еще до того, как они становятся доступными наблюдению в результате их выделения и накопления на противоположных сторонах дифференцирующихся клеток. Хотя при рассмотрении клетки под самым сильным микроскопом ее содержимое может казаться однородным, если не считать суспендированных в нем тонкозернистых гранул, вполне возможно и даже несомненно, что в клетке находятся в уже сформированном и индивидуализированном виде разнообразные физиологические молекулы. Процесс разделения, который мы видим, - это лишь следствие дифференцировки, уже существующей, но невидимой.»
Спустя сто лет после провидческой гипотезы Ланкестера мы начинаем постигать сущность «физиологических молекул», участвующих в морфогенезе. В только что оплодотворенных яйцах имеются три информационные системы, взаимодействующие во время развития: ДНК ядерного генома, распределенные по отдельным участкам цитоплазмы информационные макромолекулы и цитоскелетный матрикс, регулирующий местоположение локальных молекулярных событий в цитоплазме. Взаимодействиям между этими информационными элементами в развитии, их изменениям и роли в эволюции главных типов многоклеточных животных и посвящена настоящая глава.
Как указывал К. Бэр, в пределах любой группы животных развитие обычно консервативно. Механическая основа этого наблюдения очевидна. Родственные организмы представляют собой разнообразные выражения одного общего плана строения, основанного на наследовании общего типа развития. Для морфогенеза необходим необычайно сложный комплекс каскадно включающихся взаимодействий, происходящих в зародыше. Ранние стадии развития особенно устойчивы к эволюционным изменениям, потому что внесение любого изменения на ранних стадиях развития оказывает очень глубокое влияние на весь последующий процесс развития. Новый план строения требует значительной модификации всего типа развития; поэтому изменений ранних стадий развития следует ожидать в тех случаях, когда дивергенция организмов достигает такой степени, какую можно видеть между высшими таксономическими категориями - классами или типами. И действительно, в некоторых случаях эволюция новых групп сопровождалась радикальными преобразованиями организаций яйца и зародыша. Примечательно, однако, что ранние стадии развития в некоторых группах оказались столь консервативными, что, в то время как морфология поздних стадий развития и взрослых особей претерпевала глубокие изменения, организация яиц и их дробление упорно оставались сходными. Этот консерватизм послужил одной из главных основ для построения филогенетического древа Metazoa, представленного на рис. 4-1.
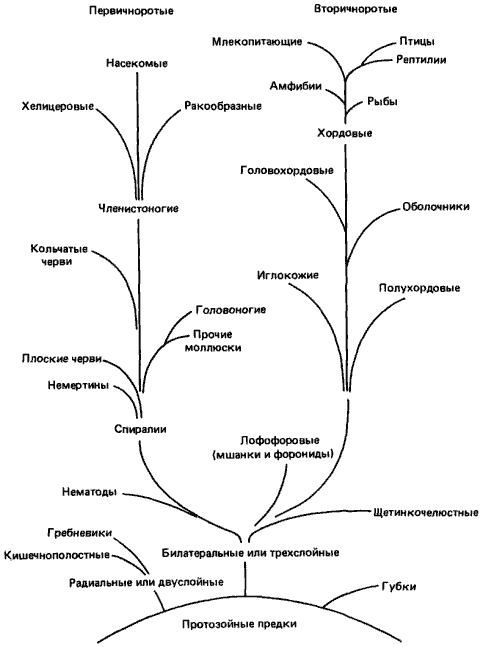
Рис. 4-1. Филогенетическое древо Metazoa.
Главный ствол филогенетического древа Metazoa делится на две большие ветви, к каждой из которых принадлежит по нескольку типов. На первый взгляд кажется нелогичным объединение таких несходных типов, как хордовые и иглокожие, в группу вторичноротых или же плоских червей, кольчецов, моллюсков и членистоногих - в группу первичноротых. Но если подробно и внимательно изучить эмбриологию этих форм, то связи между ними начинают выявляться.
Вторичноротые называются так потому, что ротовое отверстие личинки возникает несколько кпереди от бластопора, или того места, где при гаструляции происходит инвагинация клеток, из которых затем образуется первичная кишка зародыша. Яйца вторичноротых после оплодотворения начинают дробиться; при этом митотические веретена в бластомерах попеременно располагаются то параллельно, то перпендикулярно анимально-вегетативной оси яйца. Образующиеся в результате такого радиального дробления бластомеры (или клетки зародыша) располагаются точно друг над другом, как это показано на рис. 4-2 на примере яйца иглокожих. У примитивных позвоночных, таких как амфибии, дробление происходит в основном аналогичным образом.
Подобно вторичноротым, первичноротые получили свое название по месту возникновения у их личинок ротового отверстия, которое образуется у них из бластопора или вблизи него. У большинства главных типов первичноротых дробление спиральное (рис. 4-2). Все эти типы, в том числе немертины, плоские черви, моллюски и кольчецы, объединенные на рис. 4-1 в группу Spiralia, обладают на ранних стадиях эмбрионального развития очень сходным спиральным дроблением, которое, несмотря на в корне различную морфологию взрослых особей этих типов, выдает их близкое эволюционное родство.
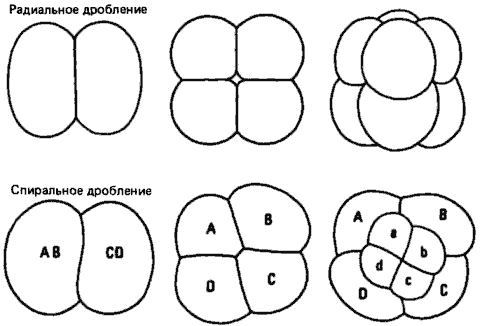
Рис. 4-2. Радиальное и спиральное дробление. Показаны зародыши на стадиях двух, четырех и восьми бластомеров. Радиальное дробление типично для иглокожих и других вторичноротых, а спиральное дробление - для многих первичноротых.
На родственные отношения указывает также другой эволюционно консервативный и фундаментальный аспект организации яйца - регионализация, т.е. распределение информационных макромолекул по определенным участкам цитоплазмы. Как у вторичноротых, так и у первичноротых зародыши содержат такие локализованные информационные системы, играющие важную роль в определении судьбы отдельных участков зародыша в процессе развития. Характер локализации этих участков тесно коррелирует с планом дробления, который в свою очередь определяется временем последовательных митотических делений и расположением веретен, что отражает организацию цитоскелетного матрикса яйца. Важнейший аспект функции и эволюции типов организации яиц заключается в том, каким образом эти типы организации обеспечивают дифференциальную экспрессию генов в клетках развивающихся зародышей, и притом в строго определенных их частях.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
2. «Метаболическая» и «структурная» гипотезы
2. «Метаболическая» и «структурная» гипотезы Группу «метаболических» гипотез составляют все те представления о природе стабильности, в которых фигурирует какое-то активирующее вещество: оно может некоторое время находиться вне ДНК, но оно должно быть способно
Глава VII Половая эволюция
Глава VII Половая эволюция Эволюция всякого живого существа происходит двояко: 1) его онтогения, или вся последовательность развития индивидуума, как такового, считая от зачатия вплоть до его естественной смерти, обусловленной старостью, и 2) его филогения, или развитие его
Глава 6 Эволюция
Глава 6 Эволюция Естественный отборУченым, который открыл научному миру эволюционный механизм, был английский натуралист Чарлз Роберт Дарвин (1809—1882), внук Эразма Дарвина, упомянутого выше.В молодости Дарвин пытался изучать медицину, а позже подумывал о посвящении в
Глава 7. Созидающая эволюция
Глава 7. Созидающая эволюция Многие люди полагают, что естественный отбор — это лишь отрицающая сила, способная к искоренению причуд и неудач, но неспособная к созиданию сложности, красоты и эффективности проекта. Действительно ли она просто отнимает от того, что уже
Глава 2. Эволюция
Глава 2. Эволюция Путь от амебы к человеку казался философам очевидным прогрессом. Хотя неизвестно, согласилась бы с этим мнением амеба. Б. Рассел (1872–1970), английский философ, лауреат Нобелевской премии 1950 г. Теория эволюции – это не только общебиологическая теория, но и
Структурная организация цепи тканевого дыхания
Структурная организация цепи тканевого дыхания Компоненты дыхательной цепи во внутренней мембране михохондрий формируют комплексы:1. I комплекс (НАДН-КоQН2-редуктаза) – принимает электороны от митохондриального НАДН и транспортирует их на КоQ. Протоны транспортируются
9.2. Эволюция и социальная организация общества
9.2. Эволюция и социальная организация общества Наш привычный мир техногенной цивилизации существует буквально одно мгновение по сравнению со временем формирования человека (в том числе и его психики) в процессе естественного отбора. Естественный отбор – медленный
Глава IV ЭВОЛЮЦИЯ ЧЕЛОВЕКА
Глава IV ЭВОЛЮЦИЯ ЧЕЛОВЕКА Так не скорби о том, что ты умер, Пелид благородный! Гомер. Одиссея. Мысль может порождать Субстанцию, и населять свои миры Созданьями, что ярче, чем живые, и давать Дыханье формам, которые всю плоть переживут. Байрон. Сон. Я начал писать эту книгу
Сравнительный обзор развития двигательной активности зародышей
Сравнительный обзор развития двигательной активности зародышей Беспозвоночные Эмбриональное поведение беспозвоночных еще очень слабо изучено. Те немногие сведения, которые пока получены, относятся в основном к кольчатым червям, моллюскам и членистоногим. Известно,
Глава VII. Эволюция людей
Глава VII. Эволюция людей 7.1. Эволюция приматов; возможные пути эволюции гоминин после их отделения от шимпанзе 7.1.1. Ранние предшественники современных людей Плацентарные млекопитающие появились по разным данным 150–100 млн лет тому назад (Kumar and Hedges, 1998; Archibald et al., 2001; Douzery et
6. Популяция как структурная единица вида
6. Популяция как структурная единица вида Вспомните!Что такое популяция?Почему организмы большинства видов живут в природе группами?Вид представляет собой сложную систему внутривидовых групп, складывающуюся в процессе эволюции в определённых условиях. Наиболее
Глава 7 Созидательная эволюция
Глава 7 Созидательная эволюция Иногда люди думают, что естественный отбор — сила исключительно отрицательная, способная только устранять уродцев и неудачников, но никак не создавать сложность, красоту и эффективность конструкции. Разве он не удаляет просто-напросто
Глава 15 Эволюция интеллекта
Глава 15 Эволюция интеллекта 1. Общие принципы эволюции интеллекта 2. Млекопитающие 3. Птицы 4. Приматы 5. Гоминиды 6. Величины IQ обезьян, человекообразных обезьян и гоминид до Homo sapiens Вернемся теперь к