Происхождение многоклеточных организмов
Происхождение многоклеточных организмов
В своей книге «Феномен человека» Тейяр де Шарден (Teilhard de Chardin) пишет по поводу одной из самых трудных загадок палеонтологической летописи - внезапного появления новых организмов: «Начальные стадии обладают досадной, но неизбежной хрупкостью, которую необходимо иметь в виду всем, кто занимается историей». Хрупкость начальных стадий и трудность их выявления, несомненно, обусловлены отчасти разрушительными воздействиями времени, постепенно уничтожающими палеонтологическую летопись. Однако становится все более ясно, что дело не только в этом. Внезапные появления новых форм - не просто артефакты. Эволюции несвойственно плавное и безмятежное течение. Скорости эволюции подвержены резким изменениям, причем многие важные и даже революционизирующие изменения в морфологии возникли за сравнительно короткое время, а, кроме того, в эволюции происходили и качественные сдвиги.
Палеонтологическая летопись первых четырех пятых истории жизни резко отличается от последней пятой ее части, в которой в изобилии содержатся остатки многоклеточных организмов. Почти на всем протяжении бесконечно долгого докембрия эволюция происходила главным образом на клеточном и биохимическом уровнях. К сожалению, у нас нет данных о самых ранних событиях, связанных с возникновением жизни и появлением наиболее примитивных организмов. Прокариотические клетки, по-видимому, уже существовали 3,4-3,0 · 109 лет назад, поскольку в породах этого возраста найдены ясные следы жизни и древнейших ископаемых бактерий.
Прокариоты - клетки, не имеющие ограниченного мембраной ядра, - господствовали на протяжении большей части докембрия. Это была эра бактерий и сине-зеленых водорослей, метаболически активных, но однообразных. Тем не менее именно от некоторых прогрессировавших прокариот произошли первые ядерные клетки - эукариоты. Самые древние эукариоты возникли, по-видимому, примерно 1,3 · 109 лет назад и были представлены по большей части простыми шарообразными и нитевидными водорослями. Однако среди них были и удивительные макроскопические формы - лентовидные водоросли, описанные Уолтером (Walter) и его сотрудниками. Определение времени возникновения эукариот на основании палеонтологической летописи затруднительно, так как критериев, с помощью которых можно было бы отличать самых ранних эукариот от прокариот, немного и в некоторых случаях они вызывают возражения. Шопф (J. W. Schopf) составил список ряда критериев, основанных на величине, форме и морфологической сложности ископаемых клеток. Среди структур, интерпретируемых как эукариоты, есть ветвящиеся нити с внутренними поперечными перегородками, сложные (например, бутылеобразные) микроископаемые, крупные цисты водорослей, клетки, содержащие плотные тельца, напоминающие остатки органелл эукариотических клеток, и тетрады клеток или спор, вероятно представляющие собой продукты мейоза. По мнению Шопфа, эти тетрады служат указанием на возникновение у эукариот пола примерно 0,9 · 109 лет назад. К сожалению, принадлежность некоторых из этих структур эукариотическим клеткам вызывает сомнение, поскольку эксперименты, проведенные Ноллом и Баргхорном (Knoll, Barghoorn) на культурах ныне живущих сине-зеленых водорослей, показали, что в дегенерирующих клетках этих водорослей появляются образования, напоминающие органеллы. Браун (Brown) и Болд (Bold), а также Элер (Oehler) и его сотрудники нашли сине-зеленые водоросли, образующие тетрады, не имеющие отношения к мейозу. При интерпретации таких трудных объектов, как ископаемые клетки, известная доля скептицизма уместна, однако мы, вероятно, имеем основания признать, что формация Биттер-Спрингс возрастом 0,9-109 лет, описанная Шопфом, содержит скопление разнообразных эукариот и что общие черты организации эукариотических клеток к этому времени уже вполне определились.
Одна из выдающихся особенностей организации эукариотической клетки - это наличие в ней окруженных мембранами органелл - митохондрий и хлоропластов, содержащих небольшие собственные ДНК-геномы и синтезирующих ограниченное число собственных белков. Эти геномы, которые имеют жизненно важное значение для сборки и функционирования органелл и для выживания клетки в целом, возникли на ранних этапах истории эукариот. Рэф (Raff) и Малер (Mahler) предложили механизм, с помощью которого они могли развиться. Главные последствия эволюции геномов у органелл эукариот заключались в том, что наличие в пределах одной клетки нескольких геномов сделало необходимой эволюцию механизмов, регулирующих и координирующих их функциональные взаимодействия. Геномы органелл контролируются ядерным геномом, и их деятельность координирована с деятельностью ядерных генов. Ядерные геномы таким образом могут взаимодействовать с другими связанными с ними геномами. Это, возможно, оказалось одной из решающих преадаптаций к развитию многоклеточной организации, которая требует координации между геномами разных клеток данного организма.
Большая часть генетических и молекулярных механизмов, необходимых для развития и дифференцировки многоклеточных организмов, возникла в процессе эволюции одноклеточных эукариот. На существование такой преадаптаций указывают многочисленные независимые попытки перехода к многоклеточности в разных группах эукариот. В своей книге «Эволюция развития» Дж. Боннер (J. Bonner) перечисляет по крайней мере десять таких попыток, результаты которых до сих пор сохранились в виде живых организмов. Проблема многоклеточности решалась по-разному. У слизевиков скопления независимых амебоидных клеток образуют многоклеточную репродуктивную фазу. Вольвокс - зеленая водоросль, состоящая из нескольких тысяч клеток, - обладает единственным в своем роде планом строения, сложным типом развития, и линия соматических клеток обособлена у нее от зародышевой линии. В процессе эволюции независимо возникли координированные и высокодифференцированные многоклеточные формы - растения, грибы, губки и животные.
Радиация многоклеточных животных впервые произошла в конце докембрийской эры. Остатки первых мягкотелых Metazoa сохранились в породах возрастом 0,7-0,6 · 109 лет в Австралии, Канаде, Англии и Южной Африке. Эта фауна получила название эдиакарской по местности в Австралии, где были обнаружены ископаемые наилучшей сохранности. Причины, вызвавшие радиацию Metazoa, и время, когда она происходила, породили множество спекуляций, поскольку фактических данных, которые сдерживали бы воображение, очень мало. Эукариоты уже существовали в течение нескольких миллионов лет, прежде чем появились первые известные нам Metazoa. Этот промежуток времени, возможно, понадобился для эволюции механизмов, необходимых для возникновения многоклеточности. Но столь же вероятно, что эти механизмы уже существовали задолго до эволюции Metazoa и что радиация последних стала в конечном счете возможной благодаря экологическим изменениям, произошедшим в конце докембрийской эры.
Одна серьезная возможность, которую выдвинули Беркнер (Berkner) и Маршалл (Marshall), состоит в том, что только в позднем докембрии содержание свободного кислорода в среде достигло такого уровня, при котором могли существовать Metazoa. Обсуждению подверглись также некоторые биохимические следствия, вытекающие из этой гипотезы. Тауе (Towe) высказал мнение, что до тех пор, пока содержание кислорода в среде не достигло достаточно высокою уровня, животные вырабатывали слишком мало коллагена, для синтеза которого необходим молекулярный кислород; поэтому тело их оставалось мягким, а размеры небольшими. Рэф и Рэф (Raff, Raff) показали, что при низком напряжении кислорода примитивные Metazoa, у которых снабжение тканей кислородом происходило путем диффузии, должны были быть ограничены в отношении своей толщины и сложности строения, а Клауд (Claud) отмечает, что животные, составляющие эдиакарскую фауну, подтверждают эту гипотезу. У некоторых червей, относящихся к эдиакарской фауне, распластанное тело занимало довольно большую площадь, будучи при этом чрезвычайно тонким и мягким. Возникновение циркуляторных систем, способных переносить кислород к тканям, стало возможным только после того, как содержание кислорода в атмосфере достигло такого уровня, при котором дыхательные белки с различным сродством к кислороду могли передавать его по цепи, такой как цепь гемоглобин-миоглобин-цитохром, найденная у многих животных. Лишь после этого могло произойти замещение эдиакарских «призраков» организмами с более массивными телом и наружным скелетом.
С другой стороны, Стенли (Stanley) указал на то, что в докембрии преобладали экосистемы с одним трофическим уровнем, отличавшиеся малым разнообразием и состоявшие главным образом из сине-зеленых водорослей. Разнообразие было ограничено, потому что несколько видов водорослей наилучшим образом использовали доступное пространство и ресурсы, исключая все другие виды. Возникновение в процессе эволюции первых растительноядных форм означало, что разнообразие уже не могло контролироваться конкурентным исключением, так что стало возможным большее разнообразие продуцентов. Это в свою очередь привело к увеличению числа ниш для растительноядных форм и создало возможность для появления плотоядных организмов, а тем самым и нескольких трофических уровней. По мнению Стенли, такими экологическими первопроходцами были простейшие, но затем появление новых экологических ниш вызвало «взрывные скорости эволюции», что привело к возникновению многоклеточности.
События, происходившие на самом деле, затеряны в прошлом, но одно, по-видимому, ясно, и это отражается в гипотезах происхождения Metazoa. Радиация многоклеточных животных началась спустя значительное время после возникновения эукариот, однако, начавшись, она происходила быстро. Возникновение разнообразных и сложных морфологии, типов развития и всех основных тканей совершилось самое большее за те 200 млн. лет, которые разделяют одноклеточных эукариот формации Биттер-Спрингс, существовавшей 0,9 · 109 лет назад, и Metazoa эдиакарской фауны, возраст которой равен 0,7-109 лет.
Эдиакарскую фауну подробно изучал Глеснер (Glaessner); она целиком состоит из бесскелетных форм, среди которых преобладают кишечнополостные и кольчецы, но содержит также очень примитивных членистоногих и, возможно, одно иглокожее. В целом в ней представлены семь классов, принадлежащих к четырем типам. Среди других Metazoa одного возраста с эдиакарской фауной можно назвать маленькую коническую раковину, возможно принадлежащую моллюску из верхнего докембрия Калифорнии, описанному Тейлором (Taylor), и два других описанных Журавлевой организма из верхнего докембрия СССР, систематическое положение которых неясно. Хотя не менее пяти типов Metazoa впервые были найдены в верхнем докембрии, обильная фауна Metazoa появилась, в сущности, в нижнекембрийских отложениях, где стали часто встречаться животные с хорошо сохраняющимися твердыми частями. Симпсон (Simpson) описал из нижнего кембрия 12 классов, относящихся к 8 типам, а позднее Стенли (Stanley) выделил в этих же типах 18 классов. К середине кембрийского периода число типов, представленных и в современной фауне, по данным Валентайна (Valentine), достигло 12. Как полагают Конвей (Conway), Морис (Morris) и Уитингтон (Whittington), некоторые другие своеобразные животные, обнаруженные в среднекембрийских сланцах Берджес в Британской Колумбии, принадлежат к 10 типам, которые целиком вымерли, возможно, вследствие неудачного плана строения, возникшего в период бурной первоначальной радиации многоклеточных животных. В кембрии появилось много очень сложно организованных животных: иглокожие; трилобиты и другие членистоногие; замковые и беззамковые брахиоподы; несколько классов моллюсков, в том числе головоногие; все они были представлены довольно разнообразными формами, а их предки установлены не были.
О быстроте, с которой происходили глубокие эволюционные изменения в ранний период возникновения Metazoa, можно судить на примере эволюции иглокожих, богатая палеонтологическая летопись которых хорошо изучена и может служить моделью для обсуждения основных проблем эволюционной теории, порождаемых этой летописью. У ныне живущих иглокожих наблюдается несколько четко различающихся планов строения, причем все они подчинены пятилучевой симметрии. Среди современных иглокожих можно назвать хорошо известных морскую звезду и морских ежей, а также менее знакомых нам голотурий и морских лилий. В настоящее время существует пять классов иглокожих, однако древние иглокожие отличались большим разнообразием.
Пауль (Paul) перечисляет 15 классов, известных из кембрийских отложений, и 19 классов - из ордовика. На рис. 2-1 показано изменение разнообразия иглокожих с течением времени. В позднем докембрии существовал один возможный их представитель - Tribranchidium. В раннем кембрии было уже четыре класса иглокожих, а к ордовику разнообразие их быстро возрастало. Затем некоторые их классы стали вымирать, по мере того как они вытеснялись экспансией других более преуспевающих классов иглокожих.
В связи с историей иглокожих в том виде, в каком она нам известна, возникают три крупные эволюционные проблемы. Первая - это отсутствие каких-либо форм, которые можно было бы считать их предками. Иглокожие появляются в палеонтологической летописи, уже обладая всеми основными характерными для этого типа признаками, которые к тому же четко выражены. Вторая проблема - отсутствие переходных форм между классами. Родственные связи, показанные на рис. 2-1 прерывистыми линиями, весьма гипотетичны. Животные, изображенные на рис. 2-2, сходны по основным чертам строения, что и позволило отнести их к одному типу, однако во всем остальном даже самые древние классы иглокожих сильно отличаются друг от друга по своей морфологии.
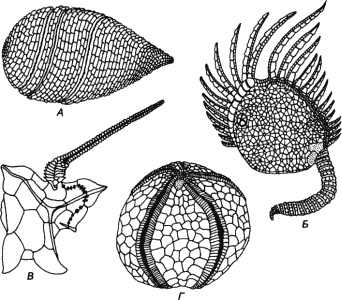
Рис. 2-2. Четыре представителя иглокожих из нижнего палеозоя (Durham, Carter, 1963; Parsley, Mintz, 1975; Ubaghs, 1971; MacBride, Spencer, 1938). А. Геликоплакоидея. Б. Паракриноидея. В. Стилофора. Г. Примитивная эхиноидея. Все эти формы были построены из кальцитных пластинок и обладали амбулакральной системой. В остальном они сильно различались как по типу симметрии, так и по общему строению. У геликоплакоидеи имелся спирально закрученный амбулакральный канал; у паракриноидеи - два таких канала, снабженные ножками; у стилофоры - один канал, проходящий по ее «руке», а у эхиноидеи - пять каналов.
Среди ранних иглокожих были классы, отличавшиеся по плану строения тела от всех ныне живущих форм. Все они построены из характерных кальцитовых пластинок и, судя по наличию у них амбулакров, обладали чрезвычайно своеобразной системой наполненных водой сосудов, имеющейся у ныне живущих иглокожих. Амбулакры служили местами прикрепления поверхностных придатков, связанных с системой сосудов и служивших для захватывания пищи, передвижения и дыхания, подобно амбулакральным ножкам ныне живущих игло кожих. Однако на этом семейное сходство кончалось. Мы привыкли к тому, что для иглокожих характерна пятилучевая симметрия, потому что все современные и большинство ископаемых форм обладают этой симметрией, однако у некоторых древних форм, таких как геликоплакоидеи, ктеноцистоидеи и паракриноидеи, в остальном несходных между собой, обнаружено асимметричное расположение частей тела, наложенное на примитивную билатеральную симметрию. Геликоплакоидеи построены из пластинок, расположенных по спирали; предполагается, что они были погружены в плотный интегумент, образуя своего рода гибкую спиральную кольчугу. Единственный раздвоенный амбулакральный канал закручен вокруг тела. Другие очень древние иглокожие также не обладают радиальной симметрией, а в некоторых случаях совершенно асимметричны. Наиболее загадочные из них это, вероятно, стилофоры. Они покрыты прочным панцирем из крупных пластинок, расположенных без всякой симметрии, и снабжены шиловидным выростом, который был, по-видимому, подвижным и который Убагс (Ubaghs) назвал аулакофором; как считает Убагс, этот вырост находился на ротовом конце животного и был связан с питанием. В отличие от этого Джефрис (Jefferies) на основании ряда особенностей внешнего и внутреннего строения стилофор считает их одним из подклассов хордовых, которому он дал название Calcichordata; у стилофор имелся сложный мозг, но они были родственны иглокожим. По мнению Джефриса, их аулакофор и амбулакр превращаются у Calcichordata в хвост с заходящей в него хордой. Гомостелии и один или два других класса также не имеют ясно выраженной симметрии. Остальные известные классы обладают радиальной симметрией, чаще всего пятилучевой. Исключение составляет известный Tribrachidium с его трехлучевой симметрией вместо пятилучевой. Эта любопытная форма и роль, которую она сыграла в изучении эволюции симметрии у иглокожих, подробно рассматривается в гл. 5.
Третья проблема касается скоростей эволюции. Эволюция ныне живущих классов иглокожих была весьма консервативной. Морские лилии, голотурии, морские звезды и морские ежи, явно сходные с существующими в настоящее время, возникли в ордовике, который окончился 450 млн. лет назад. Согласно данным в «The Fossil Record», большая часть ныне живущих отрядов морских лилий появилась в триасе и юре, так же как и большинство современных семейств правильных морских ежей, т.е. все они существуют уже почти 200 млн. лет. Несколько семейств неправильных морских ежей возникли в юре, но большая их часть моложе и появилась в меловом или третичном периоде. В других классах иглокожих имеются даже еще более древние отряды и семейства, дожившие до наших дней. Грубо говоря, третья часть семейств голотурий (класс Holothuroidea) возникла в девоне или раннем карбоне (примерно 350-400 млн. лет назад), а остальные - в юре. Единственный сохранившийся отряд из Somasteroidea существует с раннего ордовика, т.е. почти 500 млн. лет. Некоторые отряды морских звезд (класс Asteroidea) появились в ордовике, а большинство других - в ранней юре. Среди офиур (класс Ophiuroidea) есть ныне живущие подотряды, появившиеся в ордовике, силуре, девоне и юре.
И все же мы видим перед собой классы, в корне различающиеся по планам строения и возникшие уже при первой радиации иглокожих. Быть может, иглокожие имеют чрезвычайно длительную историю, не запечатленную в палеонтологической летописи и начавшуюся сотни миллионов лет назад, еще в докембрии, так что они такие же древние, как и первые ископаемые одноклеточные эукариоты. Такое предположение представляется маловероятным. Скорее, можно предположить, что первые иглокожие в позднем докембрии эволюционировали очень быстро, выработав целый спектр более или менее удачных планов строения. Последующая эволюция состояла в закреплении отдельных планов строения и возникновении разнообразия в пределах каждого из них.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
8.2. Расцвет многоклеточных организмов
8.2. Расцвет многоклеточных организмов Для дальнейших ступеней развития биотического круговорота, связанных с образованием и повсеместным распространением эукариот, более характерны процессы дифференциации и специализации. На этих этапах эволюции жизнь освоила новые
«ЗОЛОТОЙ ВЕК» МНОГОКЛЕТОЧНЫХ
«ЗОЛОТОЙ ВЕК» МНОГОКЛЕТОЧНЫХ Но загадка кембрия, загадка внезапной и мощной вспышки жизни не была решена. Миллиард лет густо зеленели у берегов водорослевые луга и пастбища. Миллиард лет тянулись к солнцу жесткие буроватые ветви подводных лесов-строматолитов.А где же
4. Формы размножения организмов
4. Формы размножения организмов Преемственность поколений организмов в природе осуществляется за счет воспроизведения. Размножение — это способность организма воспроизводить себе подобных. В природе существует два типа размножения: бесполое и половое.Виды бесполого
Происхождение многоклеточных
Происхождение многоклеточных Появление многоклеточных — типичный пример крупного эволюционного преобразования путем объединения множества простых репликаторов (в данном случае — клеток) в один комплексный.В истории земной жизни многоклеточность возникала много
11.4. Взаимоотношения организмов в природе
11.4. Взаимоотношения организмов в природе Популяционные взаимодействия определяются взаимоотношениями между отдельными особями популяций. Совокупность таких взаимоотношений и составляют действующие на популяцию биологические
Типы взаимоотношений организмов
Типы взаимоотношений организмов Существует несколько классификаций типов взаимоотношений живых организмов в природе. Можно выделить основные варианты: конкуренция, протокооперация, комменсализм, мутуализм, паразитизм, хищничество, аменсализм.Конкуренция – это
2.2. Адаптации организмов
2.2. Адаптации организмов Приспособления организмов к среде носят название адаптации. Под адаптациями понимаются любые изменения в структуре и функциях организмов, повышающие их шансы на выживание.Способность к адаптациям – одно из основных свойств жизни вообще, так как
7.3. Отношения организмов в биоценозах
7.3. Отношения организмов в биоценозах Основу возникновения и существования биоценозов представляют отношения организмов, их связи, в которые они вступают друг с другом, населяя один и тот же биотоп. Эти связи определяют основные условия жизни видов в сообществе,
22. Индивидуальное развитие организмов
22. Индивидуальное развитие организмов Вспомните!Из каких периодов складывается индивидуальное развитие организма?Что такое развитие с метаморфозом?Для каких организмов характерен такой тип развития?Индивидуальное развитие особи, всю совокупность её преобразований
4. Размножение организмов
4. Размножение организмов Организмы обладают важным биологическим свойством — способностью размножаться, т. е. воспроизводить потомство. Благодаря этой способности поддерживается и продолжается существование всех форм живых существ. Через размножение же
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни В свете современных знаний ясно, что механизмы эндотрофии и экзотрофии родственны, а не противоположны, как представлялось ранее, когда экзотрофию рассматривали в качестве гетеротрофии, а
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни
1.8. Происхождение и эволюция эндо- и экзотрофии Трофика и происхождение жизни В свете современных знаний ясно, что механизмы эндотрофии и экзотрофии родственны, а не противоположны, как представлялось ранее, когда экзотрофию рассматривали в качестве гетеротрофии, а