Гетерохрония - классический механизм эволюционных изменений
Гетерохрония - классический механизм эволюционных изменений
Попытки найти механизмы эволюции, связанные своими корнями с онтогенезом, сосредоточены главным образом на гетерохронии - изменениях относительных сроков процессов развития. Геккель подчеркивал зависимость между эволюцией и индвидуальным развитием, состоящую в том, что в онтогенезе потомка представлены в сжатом виде взрослые стадии его предков. Конечно, высказывались мнения, что эволюция может происходить в результате временных сдвигов в онтогенезе, отличных от тех, с которыми связана геккелевская рекапитуляция. Дюмериль (Dumeril) в 60-х годах прошлого века изучал аксолотля, соматическое развитие которого задержано относительно развития гонад, а другие биологи, в их числе Бэр (Baer) и Коллман (Kollman), предложивший термин неотения, описали другие примеры созревания гонад в организме личинки. В своей книге «Онтогенез и филогенез» Гулд (Gould) подробно изложил историю развития этих идей и показал, что в конце XIX в. ни один из этих случаев, явно не укладывающихся в теорию рекапитуляции, не оказал заметного воздействия на всеобщее увлечение биогенетическим законом.
Геккелевская теория рекапитуляции исходила из идеи Ламарка о том, что новые признаки могут приобретаться только у взрослых форм. Гарстанг (Garstang) своей критической статьей, опубликованной в 1922 г., решительно покончил с представлением о рекапитуляции как универсальном механизме эволюции, поскольку менделевская генетика ясно показала возможность появления и эффективного отбора признаков на любой стадии развития. Кроме того, в своих ранних исследованиях Форд (Ford) и Гексли, а также Гольдшмидт (Goldschmidt) установили способность генов регулировать скорость биологических событий. Например, Форд и Гексли в своей работе изучали скорость накопления пигмента в глазу развивающегося рачка-бокоплава (Gammarus). Они установили, что у бокоплавов, несущих идентичные аллели генов, контролирующих пигментацию глаз, окраска глаз в конечном счете оказывалась совершенно различной и зависела от того, представлен ли у них «ген скорости», регулирующий скорость накопления пигмента, доминантными или рецессивными аллелями.
Концепция генов скорости в начале 30-х годов обладала неотразимой привлекательностью для ряда биологов-эволюционистов, в число которых входили де Бер (de Beer), Гексли и Холдейн. Холдейн, например, рассматривал гены, которые, как это было установлено, действуют на разных стадиях жизненного цикла (гамета, зигота, личинка, ювенильная стадия, созревание репродуктивных органов и т.п.), и высказал мнение, что гетерохрония может быть сцеплена с изменениями в сроках действия генов, регулирующих скорости процессов развития. Поскольку гены могут действовать на любой стадии развития, глубокие эволюционные изменения морфологии, физиологических адаптации или поведения могут быть достигнуты просто за счет изменения сроков наступления тех или иных событий. Гексли, которого в это время интересовало явление аллометрии, мог объяснять относительный рост как результат действия генов скорости. Гены скорости явно могли служить средством изменения характера относительного роста в процессе эволюции. Таким образом:
«В общем ясно, что гены скорости могут мутировать либо в плюс-, либо в минус-направлении, либо ускоряя, либо замедляя процессы, на которые они воздействуют. В первом случае эффект будет, по крайней мере в некоторых отношениях, рекапитуляционным, поскольку через некое состояние, которого организм достигал прежде на взрослой стадии, он проходит теперь на более ранней стадии. Во втором случае эффект будет антирекапитуляционным, поскольку состояние, которое прежде характеризовало одну из более ранних стадий развития, теперь сместилось на взрослую стадию».
Этот новый взгляд на процессы развития как на способ эволюционного изменения создал основу для множества появляющихся в 20-е и 30-е годы филогенетических гипотез, отрицавших рекапитуляцию. Многие из этих теорий изложил де Бер (de Beer) в своей книге «Зародыши и предки». Среди них есть такие разнообразные точки зрения, как заключение Болк (Bolk) о том, что многие морфологические признаки человека - результат сохранения у взрослой особи признаков, имевшихся у зародышей ее обезьяньих предков; утверждение Гарстанга о происхождении позвоночных из головастикообразных личинок оболочников, достигших половозрелости, и предположение де Бера о подобном же происхождении насекомых из шестиногих личинок многоножек (пример, к которому мы еще вернемся). Де Бер особенно подчеркивал важность появления новых структурных признаков у не поддающихся фоссилизации личиночных стадий в качестве механизма возникновения новых групп Metazoa. Это очень соблазнительная возможность, потому что «...быть может, эти разрывы, эти прерывистости в филогенетических рядах взрослых форм до некоторой степени обусловлены „потайной“ эволюцией на ранних стадиях, за которой следовали неотения и внезапное проявление этих скрытых качественных новшеств». В этом свете некоторые из восьми типов гетерохронии, которые определяет де Бер в «Зародышах и предках», имеют очень глубокий макроэволюционный потенциал, тогда как другие типы, причем именно ведущие к рекапитуляции, дали лишь мелкие структурные модификации. Наиболее очевидные временные сдвиги связаны с диссоциацией между скоростями развития соматических признаков и скоростью созревания гонад. В сущности, классические определения категорий гетерохронии основаны на этих диссоциациях. Здесь мы придерживаемся данных Гулдом определений процессов, ведущих к рекапитуляции, и процессов, приводящих к педоморфозу. Различаемые им четыре типа гетерохронии и их результаты представлены в табл. 6-1. Если появление данного соматического признака ускоряется по сравнению с созреванием гонад, то это приведет к тому, что признак, принадлежавший прежде взрослой форме, превратится у потомков в ювенильный признак: это классический путь к геккелевской рекапитуляции. Второй путь возникновения рекапитуляции возможен в том случае, если созревание так задерживается, что признак, бывший прежде признаком взрослой формы, появляется на той же самой стадии развития, но эта стадия (в результате удлинения периода развития) оказывается теперь уже не взрослой, а «предвзрослой» стадией. Это явление носит название гиперморфоза. Увеличение размеров - обычное эволюционное направление, и оно чаще всего приводит к гиперморфозу. Ускорение соматического развития также служит способом увеличения размеров и может играть определенную роль в использовании положительных аллометрических зависимостей в эволюционных целях. Предковая морфология достигается при этом на более ранней стадии развития, а такие крайне резко выраженные структуры, как рога титанотериев или ветвистые рога Megaloceros giganteus, представляют собой следствие аллометрических зависимостей, экстраполированных на более крупные размеры тела.
Таблица 6-1. Типы гетерохронии (Gould, 1977; с небольшими изменениями)
Изменение сроков Название по системе де Бера Морфологический результат появления соматического признака созревания репродуктивных органов Ускорение Без изменения Акселерация Рекапитуляция (путем ускорения) Без изменения Ускорение Педоморфоз (прогенез) Педоморфоз (путем урезывания) Задержка Без изменения Неотения Педоморфоз (путем задержки) Без изменения Задержка Гиперморфоз Рекапитуляция (путем продления)Сдвиги сроков развития могут привести к эволюционным результатам, прямо противоположным рекапитуляции - к педоморфозу. При педоморфозе признаки, которые у предков были характерны для ювенильных стадий, сохраняются у их потомков во взрослом состоянии; это, однако, может произойти двумя совершенно различными способами. В наиболее хорошо знакомом случае - при неотении - соматическое развитие задерживается по сравнению с течением полового созревания. Например, аксолотлю - этой рабочей лошадке неотении (рис. 6-1) - требуется столько же времени для полового созревания, как и родственным ему ненеотеническим хвостатым амфибиям, но при этом многие соматические морфологические признаки остаются у него на личиночной стадии, так что половозрелый аксолотль продолжает жить в воде и выглядит как личинка-переросток. Существует и другой вид педоморфоза - прогенез, который возникает в результате совершенно иного процесса. При прогенезе созревание гонад ускоряется, так что половая зрелость достигается в маленьком и в, сущности, ювенильном теле: соматическое развитие урезано.
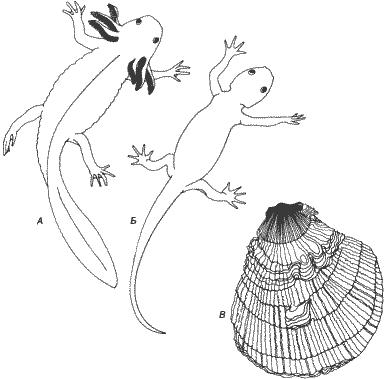
Рис. 6-1. Педоморфоз и рекапитуляция. А. Мексиканский аксолотль сохраняет жабры, уплощенный хвост и кожу, характерные для личинки, хотя он достигает размеров взрослой особи и половой зрелости. Если этой неотенической личинке вводить тироксин, то она претерпевает метаморфоз, превращаясь в типичную наземную взрослую форму (Б). В. У двустворчатого моллюска Hinnites наблюдается классическая рекапитуляция. На ювенильных стадиях Hinnites похож на других морских гребешков, но при дальнейшем созревании он прикрепляется ко дну и в результате продолжающегося роста раковины образует сходную с устрицей форму с менее упорядоченной структурой (Аксолотли с фотографий, любезно предоставленных G. M. Malacinski; Hinnites - с натуры).
Педоморфоз как одного, так и другого типа обеспечивает легкую в генетическом отношении эволюционную реакцию на давление со стороны среды, потому что при этом может быть использована морфогенетическая система, которая уже интегрирована. И неотения, и прогенез приводят к педоморфозу, но они, по-видимому, возникают в ответ на разные условия. Неотения может, например, возникнуть в ситуациях, когда неотеническая особь получает возможность избежать перехода в суровую или неустойчивую среду, в которой должен обитать взрослый организм, и остаться в более мягкой и стабильной среде, где обитают личинки. Типичным примером этого служат неотенические личинки амфибий; у некоторых групп хвостатых амфибий в процессе эволюции возникли неотенические взрослые формы, продолжающие жить в воде.
Неотения открывает довольно неожиданные эволюционные возможности, как, например, у нелетающих птиц, питающихся на земле. Неспособность к полету часто встречается у птиц, обитающих на островах, где отсутствуют хищные млекопитающие. Поскольку, как указывают Олсон (Olson) и Федуччиа (Feduccia), летательные мышцы, кости плечевого пояса и грудина составляют примерно 20-25% массы обычной птицы, то на их рост и поддержание затрачивается значительная доля ее энергетического бюджета. Если полет не дает непосредственного преимущества, то отбор не будет благоприятствовать сохранению дорогостоящего в энергетическом выражении комплекса структур. Одним из механизмов, обеспечивающих быструю утрату этих признаков, может быть неотения, и нелетающие птицы действительно во многом сходны с птенцами летающих видов. Согласно Олсону и Федуччиа, у некоторых групп, таких как голуби и пастушки, грудина развивается на поздних стадиях онтогенеза, после вылупления. Разница между ними и куриными значительна: среди куриных нет ни одного нелетающего вида. У типичного представителя куриных - курицы - окостенение грудины начинается между 8-м и 12-м днями инкубирования, а не после вылупления, как у пастушков. Задержка части морфогенетической программы на такой ранней стадии развития привела бы, возможно, к летальному исходу, однако задержка развития грудины у форм, у которых эти процессы происходят лишь после вылупления, вряд ли вызовет серьезные осложнения. Таким образом нелетающие пастушковые возникали в процессе эволюции неоднократно, потому что в программе развития пастушковых имеется нреадаптация к изменению такого типа. После того как неотеническое изменение возникло, становится возможным достижение больших размеров и другие адаптации. К сожалению, дронты, моа, эпиорнисы и все другие многочисленные виды, составлявшие великолепный ряд нелетающих птиц, населявших большие и малые острова по всему земному шару, вымерли в результате вмешательства человека в природу.
Неотенические признаки обычны у птиц и млекопитающих с высокоразвитым общественным образом жизни, и было высказано предположение, что задержанное соматическое созревание, возможно, облегчает распознавание ранга отдельных особей, позволяя избегать столкновений и повышать стабильность сообщества. Неотенические признаки человека отражают в усиленной степени такие признаки приматов, как стабильность сообщества, интенсивная забота о потомстве и длительный период созревания. В строении черепа отражены также такие Неотенические признаки, как увеличение объема головного мозга, которое было достигнуто путем увеличения продолжительности внутриутробного развития с сопровождающими его аллометрическими тенденциями.
В отличие от этого прогенез представляет собой, по-видимому, типичную реакцию на такие условия среды, в которых высокий уровень размножения или малые размеры отдельной особи могут оказаться особенно выгодными. В некоторых из таких случаев отбор может идти не на какой-то определенный морфологический признак, а на малые размеры как таковые. В результате возникает зрелый в репродуктивном отношении организм, обладающий личиночной морфологией или же смесью личиночных и взрослых признаков. В средах, открытых для небольших организмов, некоторые морфологические признаки выходят из-под ограничительных воздействий отбора, которые испытывали на себе более крупные предковые виды. В условиях ослабленного или модифицированного отбора анатомические признаки могут накапливать изменения, делающие возможным переходы от одного плана строения к другому, осуществить которые в других условиях было бы крайне трудно. Подобная точка зрения вполне приложима также к происхождению насекомых от педоморфных многоножек, как это предполагает де Бер. На рис. 6-2 изображена личинка многоножки Glomeris, у которой в момент вылупления имеется только три пары ног и ограниченное число сегментов тела. Как правило, число ног и сегментов тела увеличивается у многоножек в процессе развития. Если, однако, такая личинка достигла бы половозрелости, то получился бы организм, очень похожий на примитивное насекомое.
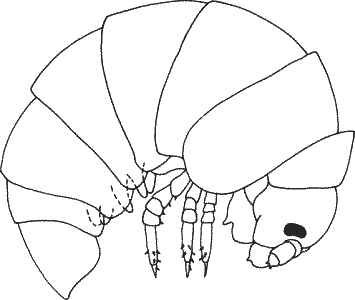
Рис. 6-2. Только что вылупившаяся личинка многоножки Glomeris с тремя парами ног (Juberthie-Jupeau, 1974).
Существенный момент здесь состоит в том, что, хотя первоначальное прогенетическое событие сохранялось отбором ввиду его непосредственного значения для выживания, оно послужило также отправной точкой для очень крупного макроэволюционного события. Возможно, что таким образом начиналась эволюция насекомых. Однако насекомые - это не просто педоморфные многоножки. Вслед за педоморфозом должен был произойти целый ряд дальнейших генетических изменений. Необходимо было приобрести не только такое качественное новшество, как крылья, но и ряд генов, регулирующих дифференцировку всех сегментов тела. В результате детерминирующего воздействия этих генов на строение сегментов развивается насекомое с иной очень стабильной дифференцировкой передне-, средне- и задне-груди и связанных с ними придатков, причем развитие конечностей на сегментах брюшка у него подавлено. Роль этих генов в развитии насекомых и их эволюции подробно обсуждается в гл. 8 и 9. Многие гетерохронии состоят в изменениях сроков появления одного соматического признака по отношению к другому при незначительном изменении остального онтогенеза. Такие гетерохронические изменения могут быть неразрывно связаны своими механизмами с изменениями индукционных взаимодействий, рассмотренных в гл. 5. Изменения сроков клеточного деления или перемещения слоев тканей должны оказывать непосредственное влияние на индукционные взаимодействия. Это может повлечь за собой увеличение размеров индуцированной структуры. И наоборот, изменение сроков может привести к утрате какой-либо индуцируемой структуры, и вовсе не потому, что индуцирующая ткань не сумела произвести индукционный сигнал, или не потому, что индуцируемая ткань утратила компетентность, необходимую для того чтобы отреагировать на этот сигнал, а потому, что этим двум тканям не удалось вступить в контакт в нужное время. Подобные примеры известны среди обитающих в пещерах позвоночных с рудиментарными глазами. Например, Шламп (Schlampp) обнаружил, что у обитающего в пещерах слепого европейского протея (Proteus anguineus) инвагинация глазного пузыря с образованием глазного бокала протекает нормально; однако преждевременная миграция мезодермы, располагающейся между глазным бокалом и эктодермой, блокирует индукцию хрусталика на ранней стадии развития. Нарушение индукции обусловливает также развитие лишь зачаточных глаз у мексиканской пещерной рыбы (Astianax). Кан (Cahn) сравнивал развитие глаз у пещерной формы и у речного вида Astianax mexicanus, от которого она произошла. В этом случае имели место два гетерохронических события: задержка образования глазных пузырей и замедление митотических делений клеток сетчатки у слепой пещерной формы по сравнению с этими процессами у речной формы. Это приводит к нарушению индукции хрусталика, а следовательно, и к развитию зачаточных глаз.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
2. Молекулярные методы изучения изменений ДНК в ядрах
2. Молекулярные методы изучения изменений ДНК в ядрах В предыдущей главе мы уже говорили о гибридизации молекул ДНК. Этот метод позволяет сравнивать, в частности, ДНК, полученные из разных стадий развития одного вида. Если гибридизация ДНК раннего зародыша и взрослого
Общие закономерности изменений клеток при их ускоренном размножении
Общие закономерности изменений клеток при их ускоренном размножении Наиболее подробно эти закономерности изложены в монографии И. Г. Акоева и Н. Н. Мотлоха «Биофизический анализ предпатологических и предлейкозных состояний» [1984]. Ниже конспективно изложены материалы из
Механизм действия ферментов
Механизм действия ферментов В любой ферментативной реакции выделяют следующую стадийность:E + S ? [ES] ?E + Pгде Е – фермент, S – субстрат, [ES] – фермент-субстратный комплекс, Р – продукт.Механизм действия ферментов может быть рассмотрен с двух позиций: с точки зрения изменения
Гетерохрония
Гетерохрония Первую классификацию явлений гетерохронии предложил Г. де Бер, выделив 8 разных типов (DeBeer G., 1958). Затем различные авторы давали другие классификации и свои термины, что создавало терминологическую путаницу (Gould S., 1977).В самом общем виде все варианты
Древесные саламандры и лягушки без головастиков. Гетерохрония и морфологическая адаптация
Древесные саламандры и лягушки без головастиков. Гетерохрония и морфологическая адаптация Почему гетерохрония может служить таким обычным способом эволюции? Ответ на этот вопрос, по-видимому, заключен в характеристике, которую дал эволюции Ф. Жакоб (F. Jacob). По его словам,
2.2. Механизм
2.2. Механизм Современная механистическая теория морфогенеза приписывает главную роль ДНК по следующим четырем причинам. Во-первых, было обнаружено, что многие случаи наследственных различий между животными или растениями данного вида зависят от генов, которые
Глава четвертая Энтропия Источник изменений
Глава четвертая Энтропия Источник изменений Не знающий второго начала термодинамики подобен тому, кто никогда не читал творений Шекспира.[14] Ч.П. Сноу Великая идея: изменения являются следствием бесцельного падения энергии и вещества в беспорядокЕсть вопрос, который
Новизна изменений
Новизна изменений Новизна прибавляет много напрасных страхов. Плутарх. Гай Марий, XVI Принципиальным отличием современного определения стресса от предложенного Селье служит указание на то, что для его развития необходима новизна изменений в окружающей среде. Отклонение
Глава 3 Накопление небольших изменений
Глава 3 Накопление небольших изменений Итак, живые существа столь невероятны и столь превосходно “продуманны”, что не могли возникнуть случайно. Тогда каким же образом они возникли? Ответ, по Дарвину, таков: путем постепенного, пошагового преобразования простейших
Глава I. Механизм эволюции
Глава I. Механизм эволюции Главная стратегическая задача биологии как научной дисциплины — познание закономерностей развития жизни для управления ими в интересах человека. Еще в 20-х годах Н. И. Вавилов поставил перед учеными проблему перехода к управляемой эволюции —
Взаимодействие эволюционных сил — механизм эволюции
Взаимодействие эволюционных сил — механизм эволюции Пусковой механизм эволюции функционирует в результате совместного действия эволюционных факторов в пределах популяции как эволюционной единицы. Всякая популяция у любого вида подвержена тому или иному давлению
Феногеография как путь выявления действия эволюционных факторов
Феногеография как путь выявления действия эволюционных факторов Среди других эволюционных факторов, кроме более подробно рассмотренного выше естественного отбора, феногеография в ряде случаев позволяет судить о действии волн численности в их ярком частном проявлении
Глава 13 Внесение изменений
Глава 13 Внесение изменений В 1821 году состоялась премьера оперы композитора Карла Марии фон Вебера «Вольный стрелок». Герой оперы, Макс, хочет жениться на Агате и, дабы произвести впечатление на отца девушки, намерен победить на состязании стрелков. Отчаянно боясь
Глава 13. Внесение изменений
Глава 13. Внесение изменений …разработал первые препараты для борьбы с сифилисом, облегчив страдания миллионов. Bosch, Rosich, 2008.…возможно, на такое название его как раз и вдохновила популярная опера Вебера. Strebhardt, Ullrich, 2008.…последнее средство, когда все иные методы лечения
Механизм компенсации дозы
Механизм компенсации дозы У подавляющего большинства млекопитающих (но не у сумчатых) в соматических клетках самок одна из Х-хромосом инактивирована. Подобное выключение является одним из вариантов решения проблемы у видов, для которых один пол представлен двумя