Скорости эволюции
Скорости эволюции
До сих пор мы рассматривали скорости эволюции вообще, не пытаясь определить, какие изменения морфологии за 1 млн. лет соответствуют горотелическим или тахителическим скоростям. Однако из палеонтологической летописи можно извлечь конкретные эволюционные данные, позволяющие в сочетании с достаточно точными измерениями абсолютного времени произвести некоторые количественные оценки скоростей эволюции. Из палеонтологической летописи можно извлечь сведения о трех обширных категориях эволюционных скоростей: скоростях таксономического изменения, скоростях изменения размеров и скоростях изменения формы. Эти скорости, конечно, не независимы друг от друга, но операционально их можно рассматривать по отдельности.
Скорости таксономического изменения наиболее субъективны, потому что отдельные систематики, занимающиеся близкими организмами, могут пользоваться разными критериями при установлении таких категорий, как роды и семейства. «Объединитель» может поместить десять близких видов в один род, а его коллега-«дробитель» отнесется к этому неодобрительно и сочтет, что они принадлежат к трем разным родам. Кроме того, и это более серьезно, систематики, работающие с сильно различающимися группами, неизбежно используют в своей работе очень разные морфологические признаки, термины и критерии. Трудно решить, имеют ли такие таксономические категории, как роды или семейства, одинаковое эволюционное значение при сравнениях организмов, принадлежащих к разным классам или типам со своими особыми планами строения и эволюционными историями. Тем не менее, не упуская из виду эти трудности, можно использовать обширные таксономические данные, добытые из палеонтологии, для оценки скоростей эволюции, особенно среди родственных организмов, или для оценки изменений скорости эволюции в пределах одной линии. Сравнения между неродственными группами более условны, однако таксономию нельзя считать совсем уж произвольной. Одни и те же общие принципы применяются исследователями всех групп, когда они пытаются создать иерархические классификации, отражающие эволюционное родство между разными группами организмов и в пределах отдельных групп.
Таким образом, таксономические категории представляют собой итог произведенных систематиком оценок степени эволюционной дивергенции и отражают уровень морфологических различий между классифицируемыми организмами. При наличии данных об абсолютных возрастах, как это иногда бывает, имеется возможность оценить время, необходимое для достижения существенных морфологических изменений. Эти оценки могут позволить нам выяснить, какая точность определения времени необходима для изучения прерывистых событий, и установить, сколь значительными могут быть горотелические скорости в действительности.
В 1953 г. Симпсон (Simpson) указал, что существует несколько способов выведения эволюционных скоростей из таксономических данных. Филогенетические скорости - это те скорости, с которыми эволюционируют таксоны, такие как виды или роды в пределах одной филогенетической линии. В идеале могут быть определены скорости для эволюционирующей линии, в которой виды или роды возникают в известное время от известных предков и исчезают не в результате вымирания, а давая начало известным группам-потомкам. Хотя достоверно датированные последовательности предков-потомков встречаются редко, известно несколько случаев, в которых возможен такой прямой подход. Эволюционный ряд аммонитов из сем. Scaphitidae, представленный на рис. 2-4, служит хорошим примером для прямого определения филогенетических скоростей. Существование отдельных видов, принадлежащих к этой линии, продолжалось от 500 000 до 1 млн лет и завершалось возникновением последующего вида. На основании данных по сходным эволюционным рядам видов позднемеловых аммонитов рода Baculites, которые изучали Гил (Gill) и Кобан (Cobban), продолжительность существования вида в среднем получила оценку 0,5 · 106 лет. Скорости возникновения видов представляют собой величины, обратные продолжительности их существования; для этих аммонитов скорость эволюции составляет 1-2 вида за 106 лет. Скорости для головоногих моллюсков близки к скоростям филогенетического изменения для сильно отличающейся от них группы животных - млекопитающих. Эволюция примитивных раннеэоценовых млекопитающих, которую изучали Гингерих и Букштейн (Gingerich и Bookstein), а также Гингерих и Клюге (Gingerich и Kluge), продолжалась около 4 млн. лет. Одна простая линия состояла из четырех видов приматов рода Pelycodus, каждый из которых существовал примерно 1 · 106 лет. Эволюция Hyopsodus (Condylarthra), происходившая в это же время, была более сложной; филогения нескольких видов этого рода была ветвящейся, и в ней имели место, по-видимому, как постепенные, так и прерывистые события. Виды Hyopsodus сохранялись на протяжении 0,3-0,7 · 106 лет, что соответствует скоростям, равным 1,5-3 вида за 106 лет. Аналогичные расчеты можно произвести для определения скоростей изменения на уровне родов; например, по оценкам Симпсона, для линии из 8 последовательных родов лошадей, существовавшей в течение примерно 60 · 106 лет, средняя скорость составляла 0,13 рода за 106 лет.
Принято считать, что аммониты и млекопитающие развивались со сходными средними скоростями, тогда как у других линий наблюдались различные, гораздо более низкие скорости изменения. Длительные сроки существования видов обнаружены у морских двустворчатых моллюсков - гребешков - от миоценовых до ныне живущих представителей рода Argopecten, изучавшихся Уоллером (Waller). Этот род существует примерно 19 · 106 лет; видообразование происходит у него как путем ветвления, так и, возможно, путем постепенной трансформации видов, образующих линейную последовательность, доходящую до настоящего времени. Средняя продолжительность существования вымерших видов Argopecten была равна примерно 5 · 106 лет, что соответствует 0,2 вида за 10б лет. Судя по этим данным, двустворчатые моллюски эволюционируют так же, как и живут, «не спеша», медленнее, чем млекопитающие.
Заметное различие в скоростях эволюции млекопитающих и двустворчатых моллюсков было подтверждено и при другом подходе к определению скоростей таксономической эволюции, основанном на кривых выживания. При этом подходе определяют среднюю продолжительность существования всех видов или других категорий в данной группе. Знания прямых эволюционных линий не требуется. Кривые выживания получают, нанося на график процент родов, которые впервые появились в какое-то данное время в прошлом и существуют до сих пор. Не удивительно, что среди родов, возникших в далеком прошлом, число родов, доживших до наших дней, меньше, чем среди родов, возникших недавно. Кривые выживания для вымерших родов получают, нанося на график процент родов, существовавших на протяжении некоторого данного периода времени, используя продолжительность времени от первого обнаружения каждого рода и до его последнего обнаружения в палеонтологической летописи. Симпсон впервые применил этот метод в 1953 г., воспользовавшись им для сравнения продолжительностей существования родов двустворчатых моллюсков и млекопитающих (рис. 2-7). На основании этих кривых Симпсон оценил продолжительность существования некоторого «усредненного» рода двустворчатых, как превосходящую в 10 раз продолжительность усредненного рода млекопитающих, и высказал мнение, что эволюция млекопитающих на уровне родов протекала в 10 раз быстрее, чем эволюция родов двустворчатых моллюсков.
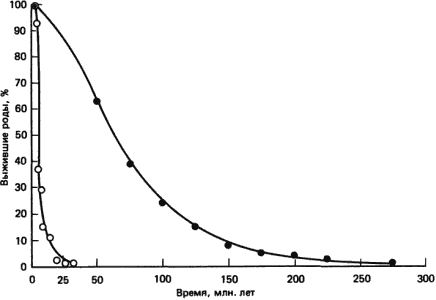
Рис. 2-7. Кривые выживания родов вымерших двустворчатых моллюсков (?) и хищных млекопитающих (?). Выживание выражено в процентах родов, просуществовавших в течение указанных по оси абсцисс периодов времени (Simpson, 1953).
Сходные результаты были получены на уровне видов. Стенли (Stanley, 1976, 1977) построил графики выживания для видов плейстоценовых млекопитающих и двустворчатых моллюсков и получил среднюю продолжительность существования для усредненного вида млекопитающих, равную примерно 1,2 · 106 лет, а для вида двустворчатых - 7 · 106 лет. Таким образом, эволюция и родов, и видов млекопитающих, как мы уже отмечали, говоря о филогенетических скоростях, протекает в 5-10 раз быстрее, чем эволюция родов и видов двустворчатых моллюсков.
Это заключение вызвало аргументированные возражения со стороны Шопфа (Schopf) и его сотрудников, которые считают, что уровень таксономического изменения, наблюдаемого в эволюционирующих линиях, может просто зависеть от их общей морфологической сложности. У более сложных организмов частей, подлежащих изменению, больше, а поэтому создается впечатление, что они эволюционируют быстрее, чем менее сложные организмы. Является ли более медленная эволюция двустворчатых по сравнению с эволюцией млекопитающих лишь кажущейся или же за их менее сложной морфологией скрывается столь же высокая скорость пока еще слабо заметной эволюции генома?
Шопф и его сотрудники подвергли это предположение проверке, оценив морфологическую сложность по числу морфологических терминов, используемых для разных групп. Как у двустворчатых моллюсков, так и у млекопитающих имеется по 3000 родов, но если для моллюсков существует всего 300 морфологических терминов, то для млекопитающих их примерно 1000. Таким образом, млекопитающие представляются морфологически более сложными, чем двустворчатые моллюски. Хотя подобная проверка согласуется с идеей о том, что скорости морфологической эволюции могут быть артефактом, обусловленным сложностью строения, тем не менее подобное объяснение явно несостоятельно. Живые ископаемые, такие как опоссум, не кажутся существенно менее сложными, чем их ближайшие родичи, эволюционирующие быстрее, а скорости эволюции могут быть выше в относительно «простых» группах, как это ясно видно по данным о скоростях эволюции, собранным Ван-Валеном (Van Valen, 1973).
Используя более свежие данные о длительности существования ископаемых организмов, чем те, которыми располагал Симпсон в 1953 г., Ван-Вален построил кривые выживания для родов и семейств многих групп. В отличие от Симпсона Ван-Вален применил логарифмическую шкалу. Оказалось, что графики выживания с использованием логарифмической шкалы более наглядны, чем графики, построенные в обычном масштабе. Если вероятность вымирания для членов данной группы остается постоянной, т. е. если один род имеет столько же шансов вымереть, как любой другой, независимо от времени их возникновения, то кривая выживания, построенная с использованием логарифмической шкалы, будет представлять собой прямую. Как показал Ван-Вален, при такой шкале график выживания действительно представляет собой прямую. На этих графиках нельзя, однако, отличить псевдовымирание (исчезновение рода в результате его эволюционного превращения в другой род) от действительного прекращения существования линии. На рис. 2-8 приведены три из графиков Ван-Валена: для родов вымерших млекопитающих, родов вымерших двустворчатых моллюсков и для родов рудистов (специализированная вымершая группа двустворчатых).
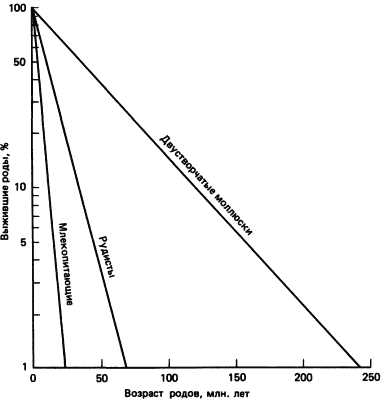
Рис. 2-8. Кривые выживания родов вымерших двустворчатых моллюсков, рудистов - специализированных мезозойских двустворчатых, принадлежащих к надсемейству Hippuritaceae, и млекопитающих (Van Valen, 1973).
Как показывает сравнение кривых для двустворчатых моллюсков и млекопитающих, период полужизни для родов первых равен 35-106 лет, т.е. средняя продолжительность существования рода равна 70 · 106 лет, тогда как значительно более крутая кривая для млекопитающих дает период полужизни, равный 3 · 106 лет, или среднюю продолжительность существования рода 6 · 106 лет. Эти данные показывают, что у млекопитающих скорость возникновения и вымирания родов примерно в 10 раз выше, чем у двустворчатых моллюсков. Совершенно очевидно, однако, что рудисты эволюционировали быстрее, чем другие двустворчатые: период полужизни составляет у них 10 · 106 лет, а средняя продолжительность существования рода 20 · 106 лет. Рудисты возникли в верхней юре и вымерли к концу мелового периода. Морфологически рудисты не были похожи на других двустворчатых моллюсков: одна створка их раковины имела форму конуса, кончик которого прикреплялся к субстрату. Другая створка служила крышечкой конуса. Некоторые виды имели огромные размеры, достигая 2 м в длину. По мнению Стенли (Stanley), большинство двустворчатых моллюсков эволюционировало медленно, так как конкуренции за ресурсы между ними не было. Рудисты жили плотными скоплениями и даже образовывали рифы. Относительно высокая скорость их эволюции, возможно, была обусловлена конкуренцией между ними за пространство. Поскольку рудисты морфологически не сложнее других двустворчатых, вряд ли следует думать, что их более быстрая эволюция представляет собой артефакт. Хотя не вызывает сомнений, что эволюция двустворчатых моллюсков в целом протекала медленно, было бы неверным считать, что они неспособны эволюционировать с высокими скоростями. Кауфман (Kauffman), изучая скорости эволюции у меловых двустворчатых моллюсков, обнаружил, что эти скорости зависят от таких факторов, как стратегия питания и уровни стресса, создаваемого условиями среды. Некоторые двустворчатые, возможно, эволюционировали так же быстро, как и млекопитающие, при такой малой средней продолжительности существования вида, как 1,25 · 106 лет. Подобно мечехвосту, двустворчатые не ограничены в отношении скорости своей эволюции какой-либо особенностью генома. Если возникает соответствующая ситуация, то геномы двустворчатых моллюсков могут отреагировать на нее, что приведет к быстрой морфологической эволюции.
По определению, скорость таксономического изменения, выведенная на основании данных по выживанию для членов какой-либо крупной группы, такой как двустворчатые моллюски, соответствует горотелии - средней скорости эволюции для рассматриваемой группы. Интересно отметить, что скорости, оцененные по данным о выживании (даже относительно высокие скорости, наблюдаемые для млекопитающих), недостаточно велики, чтобы ими можно было объяснить внезапность появления новых форм в палеонтологической летописи. Так, радиация, в ходе которой возникло большинство современных отрядов млекопитающих, происходила на протяжении палеоцена, длившегося 10-15 · 106 лет. Такую эффектную радиацию едва ли можно объяснить постепенной эволюцией в пределах линий видов со средней продолжительностью существования 1,2 · 106 лет, вычисленной Stanley для видов плейстоценовых млекопитающих.
Такой же вывод был сделан в отношении эволюции родов млекопитающих в плио-плейстоцене с применением иного подхода - определения скоростей изменения таксономических частот. В них входят скорость изменения общей частоты, например числа родов, и две скорости, определяющие общую частоту: скорость возникновения и скорость вымирания. Скорость возникновения определяется как число первых появлений в ископаемой летописи за 1 млн лет, а скорость вымирания - число последних появлений за 1 млн лет. Эти эволюционные скорости определяются легко, потому что для этого не требуется знания эволюционных линий в пределах рассматриваемой группы, а таксономическая идентификация и стратиграфическое распространение - это данные, преобладающие в палеонтологической литературе. Таким образом, для того чтобы определить скорость возникновения родов в том или ином семействе, достаточно лишь подсчитать число новых родов, появившихся в датированном стратиграфическом интервале. В 1977 г. Гингерих (Gingerich) представил данные о скоростях возникновения плио-плейстоценовых родов грызунов, парнокопытных, хищников и приматов. Во всех этих группах в течение этого времени происходила значительная радиация новых родов. Скорости возникновения были высокие: 145 родов за 106 лет у парнокопытных и 222 рода за 106 лет у грызунов. Псевдовозникновение - эволюция одного рода в другой без ветвления - позволяет объяснить только 5-20% этих скоростей. Средняя продолжительность существования одного рода грызунов равна 5,9 · 106 лет, причем половина родов сохраняется в течение примерно 2 · 106 лет. Взрывоподобную радиацию новых родов за имеющиеся для этого примерно 3 · 106 лет нельзя объяснить постепенной эволюцией. Единственная возможность совместить высокие скорости возникновения и длительное выживание - это допустить такое ветвление, при котором новые виды возникают прерывисто, а затем сохраняются на протяжении сравнительно долгого времени без дальнейших изменений.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
О течении времени, как это следует из скорости отложения осадков и размеров денудации[8] .
О течении времени, как это следует из скорости отложения осадков и размеров денудации[8]. Независимо от того, что мы не находим ископаемых остатков этих безгранично многочисленных связующих звеньев, можно, пожалуй, возразить, что не хватило бы времени на изменение
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции
Глава 1 Основы эволюции: Дарвин и синтетическая теория эволюции Пер. А. НадирянВ этой и следующей главах дается краткое описание современного состояния эволюционной биологии, какой она была до 1995 года, когда возникло новое направление науки — сравнительная геномика.
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции
Глава 2 От синтетической теории эволюции к эволюционной геномике: различные механизмы и пути эволюции Пер. А. НестеровойВ этой главе мы продолжим обсуждение эволюционной биологии в период до появления геномики. Многие из обсуждаемых направлений развития не являлись
Глава 9 Ламарковский, дарвиновский и райтовский режимы эволюции, эволюция эволюционируемости, надежность биологических систем и созидательная роль шума в эволюции
Глава 9 Ламарковский, дарвиновский и райтовский режимы эволюции, эволюция эволюционируемости, надежность биологических систем и созидательная роль шума в эволюции Пер. Д. ТулиноваДрама ламаркизмаКак уже отмечалось в предисловии к данной книге, одной из ключевых заслуг
Чему равны мировые рекорды скорости ветра?
Чему равны мировые рекорды скорости ветра? Рекордные значения скорости ветра (на высоте 10–20 метров над поверхностью Земли) составляют: в порыве – 104 метра в секунду (зафиксировано 12 апреля 1934 года на горе Вашингтон в США); средняя в течение нескольких минут – 101 метр в
Какое растение является рекордсменом по скорости роста?
Какое растение является рекордсменом по скорости роста? Самым быстрорастущим растением является бамбук (Bambusa). Скорость его роста может достигать 75 сантиметров в сутки. Рекордсменом по скорости роста среди деревьев считают эвкалипт (Eucalyptus deglupta), растущий на Новой Гвинее.
Какое животное является рекордсменом по скорости передвижения на двух ногах?
Какое животное является рекордсменом по скорости передвижения на двух ногах? Из всех животных наиболее быстро бегает на двух ногах, как ни странно, птица, хотя и не летающая. Это африканский страус, который благодаря своим мощным двупалым ногам способен поддерживать
УМЕНЬШЕНИЕ СКОРОСТИ СВЕТА В ВАКУУМЕ С 1928 по 1945 ГГ
УМЕНЬШЕНИЕ СКОРОСТИ СВЕТА В ВАКУУМЕ С 1928 по 1945 ГГ В соответствии с теорией относительности Эйнштейна скорость света в вакууме инвариантна: она является абсолютной константой. Большинство современных физических теорий основывается именно на этом постулате. Поэтому
Скорости изменения размеров
Скорости изменения размеров Изменение размеров представляет собой одно из наиболее часто встречающихся эволюционных явлений. В общем увеличение размеров - преобладающее направление эволюции. В таких различных группах, как фораминиферы и динозавры, известны примеры