Эволюция формы
Эволюция формы
Морфология реальных организмов или их частей может достигать очень высокой сложности, а поэтому производить количественные определения скоростей изменения формы труднее, чем определения скоростей таксономического изменения или изменений размеров. Это огорчительно, потому что критериями для решения таксономических проблем часто служат сложные признаки, эволюционные изменения которых могут быть очень тонкими или во всяком случае такими, что их трудно выразить в сколько-нибудь количественной форме. Скорости морфологического изменения отражаются в скоростях таксономических изменений, рассмотренных выше. Если, как считают сторонники гипотезы прерывистой эволюции, крупные эволюционные изменения всегда сопровождаются видообразованием, то в таком случае скорости таксономического изменения можно использовать для измерения темпа эволюции. Тем не менее определения скоростей таксономических изменений имеют лишь косвенное отношение к проблеме изменения формы.
Наиболее существенные морфологические изменения, происходящие в процессе эволюции, приводят к возникновению «новшеств» - новых структур, качественно отличных от существовавших прежде и открывающих возможности для новых образов жизни. Возникновение многих новшеств отражено в палеонтологической летописи. К их числу относятся конечности амфибий, яйца амниот, сочленение нижней челюсти с черепом у млекопитающих, крылья. О возникновении других новшеств, таких как гомойотермность или молочные железы, связанных с мягкими тканями или физиологическими функциями, можно судить лишь на основе изучения ныне живущих организмов. Новшество не появляется «на пустом месте». Новая структура возникает как результат изменения процессов развития предсуществующих структур. Скорости, с которыми это происходит, варьируют. Переход от сочленения нижней челюсти, типичного для рептилий, к сочленению, типичному для млекопитающих, происходил постепенно, на протяжении многих миллионов лет, и процесс этот хорошо отражен в обширных палеонтологических данных по более продвинутым звероподобным рептилиям. Что же касается скорости развития гомойотермности, то о ней можно только строить догадки. Другие новые признаки, по-видимому, развивались быстро, однако никаких данных об этом не сохранилось. Так, например, летучие мыши, которые завоевали совершенно новую для млекопитающих адаптивную зону, появляются в палеонтологической летописи внезапно, в начале эоцена. По причине скудности данных дать сколько-нибудь реальную оценку скорости, с которой происходила эта действительно глубокая реорганизация передней конечности в крыло, невозможно, однако это, по всей вероятности, имело место в период радиации плацентарных млекопитающих в палеоцене, продолжавшейся 10-12 · 106 лет.
Для эволюционных превращений таких структур, как неспециализированная передняя конечность млекопитающих, в крыло летучей мыши достаточно изменения только в программе развития, с тем чтобы те же самые компоненты расположились по-иному. Крыло летучей мыши содержит все те кости, из которых построены передние конечности других млекопитающих, и его развитие начинается с образования обычной почки конечности. Подробным рассмотрением эволюционных модификаций программ развития мы займемся в последующих главах. Здесь же нас интересует только вопрос о том, существуют ли качественные различия между эволюционными процессами, приводящими к появлению структурных новшеств, и теми процессами, в результате которых происходят более скромные эволюционные изменения морфологии. Согласно нашему рабочему допущению, различий между ними нет. Во всех эволюционных изменениях морфогенеза участвуют сходные генетические факторы, контролирующие градиенты, становление плана строения (pattern formation), скорости клеточного деления, индукционные взаимодействия и другие процессы, которые обеспечивают развитие дифференцированной структуры во всех ее деталях. Палеонтологическая летопись дает нам сведения только о двух изменениях морфологии, которые возникают в результате эволюционного изменения генов, контролирующих эти процессы, и которые можно оценить количественно. Это изменения так называемых меристических признаков и изменения аллометрических соотношений.
Меристическими называют признаки, представленные рядом идентичных или сходных структур, как, например, амбулакры иглокожих, фасетки глаза у трилобитов подсемейства Phacopinae, щетинки насекомых, ребра на раковинах брахиопод и моллюсков или позвонки и пальцы у позвоночных. Скорости изменения числа этих структур также можно выразить в единицах дарвин, предложенных Холдейном (Haldane). Ван-Вален (Van Valen) приводит различные скорости изменения меристических признаков, вычисленные на основании палеонтологических данных. Скорости изменения числа камер у некоторых ископаемых фораминифер варьируют от 70 до 120 миллидарвин. Число ребер на раковинах брахиопод изменялось со скоростями от 0 до 100 миллидарвин, а число ребер на раковинах гребешка - со скоростями от 6 до 190 миллидарвин. В 1973 г. Маглио (Maglio) вычислил в дарвинах скорости для нескольких поддающихся измерению меристических признаков зубов слонов. Изменение числа эмалевых гребней или поперечных пластин, образующих перетирающие поверхности коренных зубов слона, представлено на рис. 2-9. Показаны три линии, берущие начало от Primelephas gomphotheroides, у которого были примитивные коренные зубы с небольшим числом пластин. У рода Loxodonta, к которому принадлежит ныне живущий африканский слон, число пластин возрастало медленно, тогда как у рода Elephas, представленного в настоящее время индийским слоном, и у мамонтов (Mammuthus) оно возрастало быстро. У Elephas средняя скорость увеличения числа пластин составляла примерно 200 миллидарвин, а у поздних мамонтов она дала вспышку, достигнув 600 миллидарвин. В обеих линиях число пластин возросло от 7 до 23 примерно за 5,5 · 106 лет. В сочетании с целым набором других изменений (толщины эмали, высоты коронок и формы пластин) увеличение числа пластин привело к развитию высокоэффективных перетирающих коренных зубов.
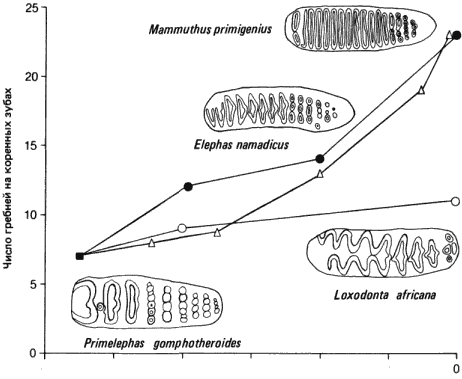
Рис. 2-9. Эволюция эмалевых гребней коренных зубов в трех линиях слонов: у индийского слона Elephas (?), африканского слона Loxodonta (?) и у мамонта Mammuthus (?) (по Maglio, 1973).
Обычно одно только изменение числа сходных частей приводит лишь к небольшим изменениям морфологии. Меньшее или большее число щетинок у мух или ребер на раковинах брахиопод не относится к категории глубоких эволюционных событий. Но в некоторых случаях изменения меристических признаков играли ключевую роль в эволюции; ярким примером служат змеи, у которых может быть до 400 позвонков и почти столько же пар ребер, что обеспечивает эффективный, хотя и весьма специализированный, способ передвижения, а в случае питонов и удавов - единственный в своем роде способ расправиться с жертвой. До сих пор мы рассматривали изменения размеров, как если бы они происходили независимо от изменений формы. Между тем такие случаи редки. Крупные животные - это обычно не просто увеличенные варианты своих более мелких предков, а взрослые особи - это не просто увеличенные молодые. Значительная часть изменений формы, происходящих в период роста данной особи или в процессе эволюции данной линии, осуществляется путем изменения относительных размеров частей тела. В обоих случаях такие модификации пропорций представляют собой результат изменений относительного роста разных частей тела в процессе развития. Это так называемая аллометрия. Рассмотрение аллометрических зависимостей дает возможность установить, какие изменения формы вызываются неравномерным ростом, а какие являются результатом изменений программы развития. Существуют аллометрические ряды трех типов:
1) ряды, образованные в результате измерения роста в процессе онтогенеза данного вида;
2) ряды, образованные близкими видами, различающимися по величине;
3) ряды, образуемые видами, составляющими одну линию в процессе эволюции.
Аллометрические зависимости между размерами двух структур часто описываются простой формулой, предложенной Гексли в 1932 г.:
y = bx?
где y - размер какой-либо одной структуры, а х - размеры всего тела или другой структуры, с которой структура у сравнивается. Член b - скалярный множитель, а ? - отношение удельных скоростей роста структур у и х. Это уравнение можно переписать в виде
log y = log b + ? log x.
Величины х и у обычно откладывают в логарифмическом масштабе по обеим осям. При этом получается линейный график с наклоном ? и с точкой пересечения с осью х, равной log b. В тех случаях, когда ? = 1, относительные размеры структур, представленных величинами х и у, постоянны независимо от их абсолютных размеров; иными словами, соотношение этих структур не изменяется, т.е. их рост происходит изометрически. Изометрический рост - это частный случай более общего спектра аллометрических зависимостей. В большинстве случаев ? ? 1 и пропорции изменяются с изменением размеров.
Один особенно интересный пример аллометрии в ходе развития обсуждается Гексли в 1932 г. в его книге «Проблемы относительного роста» и представлен здесь на рис. 2-10. У некоторых видов муравьев рабочие особи полиморфны, причем самые крупные рабочие, у которых головы и челюсти чрезмерно велики, несут функции солдат. Вероятно, муравьиной семье выгодно иметь разнообразные типы рабочих особей, каждый из которых более способен к выполнению определенного круга задач. Такой ряд рабочих особей изображен на рис. 2-10, где приведен также график отношения размера головы (х) к размерам туловища (у) для рабочих особей одного вида муравьев. В пределах вида эти отношения для рабочих различных размеров укладываются в одну аллометрическую кривую. Это означает, что, хотя более крупные рабочие выглядят иначе, чем мелкие, из-за своих огромных голов и челюстей, весь этот ряд в целом отражает проявление одного генетически детерминированного закона роста.
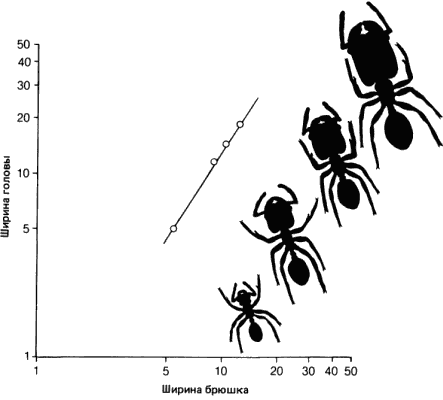
Рис. 2-10. Аллометрическая зависимость между размерами головы и тела у муравья Pheidole instabilis (Huxley, 1932; с изменениями).
До сих пор мы рассматривали только аллометрические зависимости, выявляющиеся у отдельных особей в процессе роста. Можно, однако, построить также аллометрические кривые, чтобы сравнить взрослых особей последовательных видов, образующих одну эволюционную линию. Увеличение размеров в процессе эволюции происходит, как мы убедились, очень часто. В некоторых случаях увеличение размеров имеет очень интересные последствия. Когда аллометрические тенденции, характерные для предкового вида, сохраняются, то, если для какого-либо признака ? ? 1, возникает парадоксальный результат: сохраняя тип роста, характерный для предкового вида, вид-потомок, превосходя предковый вид по размерам, приобретает иную форму. Такой способ, достойный «Алисы в стране чудес», лежит в основе морфологических изменений, наблюдаемых в некоторых хорошо известных эволюционных линиях. Одна из таких линий - травоядные млекопитающие титанотерии - достигла кульминации в олигоцене, где они представлены очень крупными формами; нос у этих форм был украшен парой массивных тупых рогов, достигавших в длину половины длины черепа. Эоценовые предки титанотериев были мельче, и рога у них либо отсутствовали, либо были короче. Размеры рогов у титанотериев радикально увеличились с увеличением общих размеров тела в процессе их эволюции в течение олигоцена. Означает ли эволюция этих крупных рогов приобретение нового варианта морфогенеза? В 1934 г. Герш (Hersh) ответил на этот вопрос отрицательно, что было неожиданным. При построении графика в логарифмическом масштабе по обеим осям все виды титанотериев укладываются в одну и ту же аллометрическую кривую с очень высоким значением ?. Следовательно, регуляция роста в процессе онтогенеза титанотерия происходила таким образом, что увеличение размеров черепа сопровождалось еще большим увеличением его носовой области. Очевидно, крупные размеры всего тела и рогов создавали селективное преимущество, и поэтому данная аллометрическая тенденция сохранялась. Аналогичным примером служат гигантские рога вымершего плейстоценового оленя Megaloceros giganteus. У крупных самцов размах рогов достигал 3-3,5 м. Гулд (Gould) показал, что рога этого оленя подчиняются тем же законам аллометрического роста, что и рога других оленей. Поскольку это был очень крупный олень, следовало ожидать, что и рога у него будут особенно большими. Но было ли это единственной причиной гигантских размеров рогов? Гулд высказал мнение, что отбор сильно благоприятствовал таким огромным рогам, поскольку они играли важную роль в брачном поведении.
Другое, более раннее, объяснение состояло в том, что эволюция рогов у титанотериев и у Megaloceros giganteus - результат ортогенеза. Согласно теории ортогенеза, несущей в себе некоторые элементы греческой трагедии, эволюция той или иной линии канализируется в определенном направлении, отклониться от которого она не может даже тогда, когда это направление перестает быть адаптивным; следовательно, ее вымирание неизбежно. Подобные представления подразумевают направленную эволюцию телеологического толка и носят мистический характер; они мало что могут дать для понимания действительных эволюционных процессов. Тем не менее в эволюции все же существуют некие направления и они должны быть исследованы.
Изменения аллометрических зависимостей в процессе эволюции могут происходить другими, возможно, более важными способами. В эволюционирующей линии может произойти изменение любого из двух параметров, ? или b. Изменение ? ведет к модификации пропорций организма в результате изменения характера роста в процессе развития. На рис. 2-11 показана аллометрическая зависимость между длиной замка и периметром раковины в линии ископаемых двустворчатых моллюсков рода Myalina, у которых происходило постепенное увеличение размеров. Точки для самых древних видов располагаются на одной кривой, более поздние - на другой, которой соответствует иное, более высокое, значение ? (больший угол наклона), и ряд завершается крупным моллюском, резко отличающимся по форме раковины от предковых видов. Все эти события в целом заняли примерно 50 · 106 лет, но время, в течение которого произошли изменения аллометрии, не превышает 10 · 106 лет.
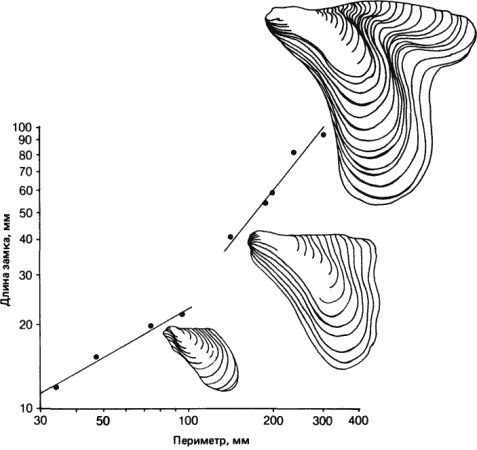
Рис. 2-11. Сдвиг аллометрической зависимости между длиной замка и периметром раковины в эволюции одной линии ископаемых двустворчатых моллюсков рода Myalina (Newell, 1942, 1949; с изменениями). В этой линии прослеживается постепенное увеличение размеров и изменение аллометрии, происходившее в течение пенсильванского и пермского периодов (см. текст).
Аналогичное изменение аллометрической зависимости между массой головного мозга и массой всего тела выявили Пилбим (Pilbeam) и Гулд (Gould) в эволюции человека. У человекообразных обезьян и у наших вымерших родичей-гоминид, принадлежавших к роду Australopithecus, значение ? равно 0,34, тогда как у ныне живущих и вымерших представителей рода Homo оно равно 1,73. Благодаря недавним открытиям Джохансона (Johanson) и Лики (Leakeys) в Восточной Африке, которые указывают на то, что дивергенция Homo от Australopithecus произошла примерно 3,5 · 106 лет назад, по мере нахождения дополнительных ископаемых остатков, относящихся к этому периоду, возможно, удастся определить, с какой скоростью происходил этот чрезвычайно важный сдвиг в аллометрии.
Значение ? для аллометрического роста, происходящего в процессе онтогенеза, часто отличается от его значений, получаемых при построении аллометрической кривой для группы взрослых форм, относящихся к родственным видам. Подобная ситуация изображена на рис. 2-12. Прерывистая линия выражает аллометрическую зависимость между массой головного мозга и массой тела для группы родственных видов насекомоядных средних дефинитивных размеров. Сплошными линиями изображены изменения аллометрических зависимостей у каждого вида на протяжении онтогенеза. В данном примере эти онтогенетические кривые имеют меньшие наклоны, чем кривая для группы видов, однако столь же вероятна и обратная картина, потому что в процессе развития значения ? часто бывают больше 1. Обратите внимание, что ?, характерное для онтогенеза, отличается от ? при сравнении взрослых особей разных видов, но при этом одинаково для всех этих видов. Однако значения b для всех видов различны.
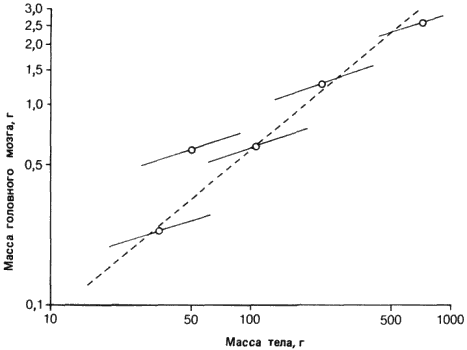
Рис. 2-12. Аллометрическая зависимость между массой головного мозга и массой тела у близких видов насекомоядных млекопитающих с Мадагаскара (прерывистая линия). Сплошными линиями показаны аллометрические зависимости в пределах каждого вида в процессе онтогенеза (Gould, 1971).
Если межвидовое ? = 1, то в таком случае взрослые особи более крупных видов представляют собой увеличенные варианты своих более мелких родичей (или предков). Гулд (Gould) высказал мнение, что это может произойти в процессе эволюции, если более крупный вид-потомок сохраняет значение ? своих мелких предков, но аллометрический рост рассматриваемой структуры начинается у него из более крупного зачатка, т.е. при более высоком значении b. Тогда, для того чтобы произошло изменение величины структуры, необходим либо сдвиг начала роста зачатка на более ранние сроки (акцелерация), либо его задержка. Это создает альтернативную возможность увеличения размеров в процессе эволюции, не требующую изменения законов роста. Если у организмов сохраняется кривая онтогенетического роста предков, а аллометрия роста сильно отличается от ? = 1, то пропорции тела могут резко изменяться с увеличением размеров. Сохраняя аллометрию, но начиная рост с других размеров зачатка, организм может избежать больших изменений в соотношении размеров разных частей.
В настоящее время все еще нет возможности полностью объяснить механизмы регуляторных процессов, лежащих в основе изометрического или аллометрического роста, однако уже начинают выявляться некоторые интересные аспекты этих процессов. Как указывает Госс (Goss), для родственных между собой организмов, сильно различающихся по общим размерам, существуют две возможности: до тех пор пока основной план их организации остается сходным, должно происходить изменение либо размеров, либо числа составных частей. По общему правилу мелкие функциональные единицы - клетки какого-либо органа или фасетки сложного глаза-изменяются в числе, тогда как крупные функциональные единицы - внутренние органы, конечности или глаза - изменяются по размеру. В известных пределах увеличение размеров приводит к повышению эффективности органов. Примером такого рода служит головной мозг: более крупные общие размеры создают возможность для большего числа нейронов и большего числа связей между ними, что приводит к усилению функционального потенциала мозга. Из этого вытекает важное следствие, состоящее в том, что в период роста соотношение размеров разных частей тела в значительной степени зависит от относительных скоростей клеточного деления. Данные о существовании факторов, регулирующих рост, появились еще в 20-е и 30-е годы; обзор ряда проведенных в этот период изящных экспериментов с пересадками органов сделал Твитти (Twitty, 1940). Так, например, при пересадках глаз от более старых аксолотлей (Ambystoma) молодым наблюдалась задержка роста этих крупных глаз, а при пересадке мелких глаз от более молодых особей более старым рост их ускорялся, так что в конечном счете величина глаз вполне соответствовала размерам их хозяина.
Существует, очевидно, какая-то система, действующая по принципу обратной связи и состоящая из циркулирующих в организме регуляторных веществ двух типов - стимулирующих клеточное деление в определенных тканях и подавляющих его. Одним из таких хорошо известных тканеспецифичных регуляторных веществ стимуляторного типа служит эритропоэтин, вырабатываемый в почках в ответ на потери крови и стимулирующий образование эритроцитов. Существуют также позитивные регуляторы, участвующие в развитии морфологических признаков. Андрогены и эстрогены, которые организм человека начинает продуцировать в подростковом возрасте, взаимодействуя с тканями-мишенями, обусловливают развитие таких хорошо заметных морфологических признаков, как вторичные половые признаки: грудь, борода и пропорции тела, характерные для взрослого человека. Эти регуляторные вещества гуморальной природы и действуют на ткани-мишени, находящиеся на расстоянии от того места, где гормоны образуются. Существуют, однако, позитивные регуляторы другого класса, которые стимулируют рост, а нередко также дифференцировку и формообразовательные процессы в ткани, непосредственно примыкающей к той ткани, которая их вырабатывает. Они обеспечивают классическую эмбриологическую индукцию, рассматриваемую в гл. 5.
Существуют также специфические ингибиторы клеточного деления, которые Баллог (Bullough) назвал кейлонами, однако они все еще недостаточно хорошо изучены. Эти вещества вырабатываются в самой ткани-мишени и подавляют ее же рост. Экспериментальное удаление или повреждение части органа, такого как печень, снижает уровень специфичного кейлона в крови и индуцирует компенсаторный рост. Как и в системе гормона роста, различные элементы этих регуляторных систем - образование регуляторов, их структура, число и специфичность рецепторных молекул - могут подвергнуться генетическому изменению в процессе эволюции новых морфологических пропорций.
Изменения в аллометрии могут происходить довольно постепенно, как это установил Кертен (Kurten) для гиен и других плейстоценовых млекопитающих, или очень быстро, как при выведении разных пород собак, что было сделано за несколько столетий. Сильная борзая, выведенная для охоты на волков, и пекинес с толстым приплюснутым носом, выведенный специально для того (к вящему удивлению), чтобы его можно было держать на коленях, различаются по величине и пропорциям. У собак обнаружены гены, детерминирующие такие признаки, как длина ног, длина морды и общие размеры. Эти признаки широко использовались селекционерами-кинологами. Высокие скорости изменений аллометрии известны также для линий, эволюционирующих в природных условиях, например у некоторых гавайских Drosophila (см. гл. 3).
Сведения о механизмах эволюции, которые можно почерпнуть из палеонтологической летописи, весьма ограничены. Сохранение ископаемых остатков какого-либо вида на всем протяжении его ареала в течение достаточно длительного периода его существования, с тем чтобы можно было застать периферические изоляты в момент акта видообразования, нельзя считать невероятным, однако число таких примеров будет ничтожным. Недоступны нам и генетические системы вымерших организмов, хотя некоторое представление о них, несомненно, дает изучение их ныне живущих родичей. Онтогенезы некоторых ископаемых организмов хорошо известны: личиночные стадии нескольких трилобитов, рост спирально закрученных раковин аммонитов и, наконец, яйца, молодые и взрослые особи знаменитого монгольского динозавра Protoceratops. Есть и другие примеры, но в целом палеонтологические «ясли» нельзя назвать переполненными. Палеонтологическая летопись свидетельствует о том, что скорости эволюции сильно варьируют независимо от того, оцениваем ли мы их по таксономическим изменениям, изменениям размеров или аллометрии. Это, пожалуй, один из главных вкладов палеобиологии в рассматриваемую здесь проблему, не говоря уже о тех поразительных свидетельствах о жизни в прошлом и о множестве исчезнувших миров, которые она дает. Мы освободились от концепции эволюции путем создания новых генов в результате постепенного замещения нуклеотидов и вынуждены искать механизмы эволюции на уровне организации генов и их экспрессии в процессе онтогенеза, с тем чтобы объяснить быстрые и глубокие изменения морфологии.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
4. Формы размножения организмов
4. Формы размножения организмов Преемственность поколений организмов в природе осуществляется за счет воспроизведения. Размножение — это способность организма воспроизводить себе подобных. В природе существует два типа размножения: бесполое и половое.Виды бесполого
3. Формы иммунного ответа
3. Формы иммунного ответа Иммунный ответ – это цепь последовательных сложных кооперативных процессов, идущих в иммунной системе в ответ на действие антигена в организме.Различают:1) первичный иммунный ответ (возникает при первой встрече с антигеном);2) вторичный
Формы перехода.
Формы перехода. Если бы возможно было показать, что существует сложный орган, который не мог образоваться путем многочисленных последовательных слабых модификаций, моя теория потерпела бы полное крушение. Но я не могу найти такого случая. Без сомнения, существуют
Пресноводные формы.
Пресноводные формы. Так как озера и речные системы отделены друг от друга участками суши, то можно бы думать, что пресноводные формы не бывают широко распространены в пределах одной страны, а так как море представляет собою еще более крупную преграду, то они никогда не
Миграции: эволюция размеров и формы тела
Миграции: эволюция размеров и формы тела Как я полагаю, предки современных сторожевых пастушьих собак появлялись в разных районах мира вместе с древними кочевниками. Так, македонские собаки, вероятно, пришли в Македонию с турецкими пастухами-мусульманами из Анатолии, а
3.1. Проблема формы
3.1. Проблема формы Не очевидно, что форма вообще представляет какую-то проблему. Окружающий нас мир полон форм, мы узнаем их в каждом акте восприятия. Но легко забываем, что существует глубокая пропасть между этим аспектом нашего опыта, который мы воспринимаем просто как
Глава 7. Наследование формы
Глава 7. Наследование формы 7.1. Генетика и наследственность Наследственные различия между организмами, одинаковыми в других отношениях, зависят от генетических различий; эти последние зависят от различий в структуре ДНК или в ее расположении в хромосомах, а эти различия
Три формы общественной жизни
Три формы общественной жизни Белая жидкостьНеобычная погода для начала августа! Гор не видно. Они скрыты облаками. Тучи нависли над городом и вот-вот опустятся еще ниже и закроют верхушки высоких тополей и многоэтажных зданий. Вместо нещадного жаркого солнца — сырость,
Формы мышления
Формы мышления На основе многолетних экспериментальных исследований Ладыгина-Котс пришла к выводу, что мышление животных имеет всегда конкретный чувственно-двигательный характер, что это мышление в действии, причем эти действия являются всегда предметно-отнесенными.
Формы смещенной активности
Формы смещенной активности Смещенная активность имеет одну из трех следующих форм:Мозаичная активность. Мозаичным называется поведение, состоящее из фрагментов различных ФКД. Герой рассказа А. П. Чехова «Папаша»[104], застигнутый женой наедине с горничной, «скинул сюртук
ЖИЗНЕННЫЕ ФОРМЫ
ЖИЗНЕННЫЕ ФОРМЫ Пожалуй, любой человек, не обладающий специальными знаниями в ботанике, может дать приблизительное описание большинства мировых биомов: в тропических дождевых лесах много деревьев; в степях и саваннах мало или нет деревьев, но много травы; в пустынях
3.4. Различные формы праны
3.4. Различные формы праны Прана -это собирательное определение пяти индивидуальных функций праны, таких как ветры (по-санскритски Ваю) в человеческом теле. Они таковы: Прана, Апаш, Самана, Удана и Въяна, в связи с чем прана выступает и как собирательная, и как индивидуальная