Морфогенетические взаимодействия и макроэволюция
Морфогенетические взаимодействия и макроэволюция
Приведенные выше примеры, относящиеся к амфибиям и птицам, иллюстрируют эволюционные последствия изменений, вызванных взаимодействиями на относительно поздних стадиях развития. Соответственно достигаемые при этом изменения представляют собой модификации уже сложившегося плана строения, а не новое направление. Это не умаляет роли таких новшеств, какие были только что описаны для покровов тела; они оказали глубокое воздействие на физиологические и поведенческие адаптации обладающих ими организмов. Но мы сталкиваемся с реальными трудностями, когда пытаемся найти объяснение относительно крупным эволюционным изменениям, приведшим к возникновению новых структур или даже совершенно новых планов строения. Такие макроэволюционные события кажутся необходимыми для того, чтобы объяснить быструю начальную радиацию типов Metazoa или возникновение таких отклоняющихся групп, как летучие мыши, появляющиеся в палеонтологической летописи с внезапностью, вызывающей тревогу. Если значительные изменения в процессах онтогенеза происходят на ранних стадиях развития, они могут привести к крупным изменениям плана строения. Конечно, при этом высока вероятность того, что такие изменения вызовут резкие нарушения онтогенеза, результатом которых окажутся нежизнеспособные зародыши. Тем не менее притягательная сила «перспективных монстров» (термин, который предложил Рихард Гольдшмидт для обозначения глубоких эволюционных изменений, достигаемых таким образом) очень велика, потому что в тех редких случаях, когда такие монстры оказываются жизнеспособными, они создают основу для объяснения макроэволюции. Неудивительно, что концепция «перспективных монстров» вызвала неистовую оппозицию не только ввиду сомнительности предложенных для ее объяснения генетических механизмов, но также и потому, что она в корне противоречила идее постепенной эволюции, основанной на популяционно-генетической теории замещения аллелей в популяциях. Например, Симпсон пишет в своей автобиографии о том, какое сильное впечатление произвела на него книга Добржанского «Генетика и происхождение видов», опубликованная в 1937 г. и побудившая его начать работу над «Темпами и формами эволюции», в которых он намеревался объединить палеонтологию с генетическим подходом к эволюции. В то же время идеи Шиндевольфа и Гольдшмидта, которые занимались поисками крупных мутаций, затрагивающих организацию генома и изменяющих раннее развитие, стимулировали Симпсона «в меньшей степени и в прямо противоположном направлении». Идеи Шиндевольфа и Гольдшмидта были чересчур смелыми, и такая реакция на них была оправданна. Очень жаль, однако, что красочный и емкий термин «перспективные монстры» породил резко отрицательное отношение к основному тезису Гольдшмидта о том, что мутациям, оказывающим влияние на развитие, принадлежит центральное место в эволюции.
Макроэволюционные изменения в развитии не обязательно должны быть экстремальными. Мы полагаем, что первые шаги для быстрых и в итоге существенных эволюционных изменений могут вызываться нелетальными генетическими нарушениями функций небольшого числа ключевых генов, играющих роль регуляторных. Начальные «легкие» генетические изменения, которые могут вызвать существенные преобразования в организме и закрепиться в небольшой популяции, должны быть жизнеспособны и открывать широкие возможности для отбора последовательных генетических изменений. Таким путем могут быстро возникать глубокие изменения, без всяких мгновенно появляющихся «перспективных монстров». Как уже ясно показала эволюция гавайских Drosophilidae, рассмотренная в гл. 3, изменения небольшого числа генов могут быть причиной резких морфологических изменений. Дальнейшие генетические данные о числе и действии регуляторных генов излагаются в гл. 8.
Существуют, по-видимому, два главных способа, для того чтобы вызвать во взаимодействиях между тканями зародыша такие изменения, которые могут привести к макроэволюционным изменениям. Следует помнить, что это не единственные способы; другим, не менее важным механизмом служит гетерохрония, рассматриваемая в гл. 6. Крупномасштабные изменения во взаимодействиях могут происходить в результате либо возрождения прежних типов интеграции, но на новых местах, либо возникновения новых взаимодействий. Мы полагаем, что действие первого из этих механизмов можно видеть в способе приобретения наружной раковины пелагическим осьминогом Argonauta, а второго - в происхождении пятилучевой симметрии у иглокожих.
У самых древних головоногих были наружные раковины. По мере роста животное перемещалось вперед по своей раковине, отгораживая одну за другой заполненные воздухом камеры, которые обеспечивают плавучесть, уравновешивая массу раковины и внутренних органов. Ныне живущие виды Nautilus - последние остатки когда-то многочисленной группы животных, вытесненной теперь более знакомыми нам головоногими сем. Coleoidae, кальмарами и осьминогами. Coleoidae совершенно утратили наружную раковину и компенсировали эту утрату поведенческими адаптациями, в том числе более совершенной координацией, большей способностью к плаванию, более развитыми органами чувств и «смышленностью». Кальмары и каракатицы сохраняют внутреннюю раковину или ее зачаток, а осьминоги утратили даже такой зачаток.
Таким образом возникает любопытная ситуация: у небольшой группы пелагических осьминогов рода Agonauta самка (рис. 5-11) обладает раковиной, очень сходной по внешней форме с раковиной аммонитов или Nautiloidae. Эта раковина отличается от раковины древних головоногих двумя существенными особенностями: она не разделена на камеры и секретируется не мантией, а парой специализированных рук. Концы этих рук сильно расширены, образуя широкие перепончатые придатки. Во время секреции раковины животное держит руки над телом и имеющиеся в перепонке железы секретируют кальцитную раковину. Две половинки раковины соединяются, образуя киль, покрытый бугорками, которые, по-видимому, соответствуют местоположению имеющихся на руках присосок. Раковина аргонавта не могла произойти от раковины древних головоногих. Это - конвергентная структура, независимо приобретенная одним из членов группы, утратившей прежнюю раковину, так что ее следует считать новшеством.
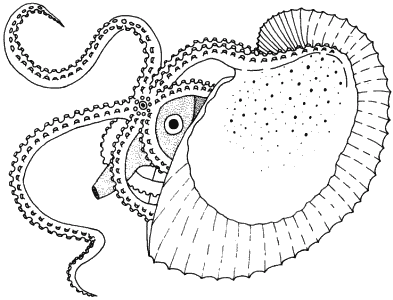
Рис. 5-11. Самка аргонавта Argonauta argo; раковина прикрепляется при помощи особых выростов, имеющихся на двух специализированных руках (Young, 1959-1960; с изменениями).
Раковина аргонавта должна давать приют как массе яиц, откладываемых самкой, так и самой самке, а поэтому представляется вероятным, что эта раковина возникла у предков аргонавта в виде простой необызвествленной оболочки для яиц. Оболочка секретируется железами, расположенными на поверхности рук. Образование известковой раковины стало возможным благодаря развитию в добавление к железам, уже имеющимся в коже, желез, секретирующих углекислый кальций. Из какого источника могли возникнуть такие раковинные железы? Они могли возникнуть de novo, или, что более вероятно, у аргонавта произошла реактивация древней и давно подавленной морфогенетической программы дифференцировки раковинных желез. Вместо того чтобы развиваться в мантии, где они располагались у предковых форм, эти железы образуются на руках. Подавление прежних типов экспрессии генов, контролирующих морфогенез, возможно, происходит гораздо раньше, чем утрачиваются сами гены. Так, несомненно, обстояло дело с утратой зубов курами; в этом случае с тех пор, когда у предков современных птиц клюв был снабжен зубами, прошло почти 100 млн. лет. Если гены, связанные с данным признаком, участвуют в других индукционных событиях, они могут вообще не утрачиваться, хотя они уже не экспрессируются с образованием предковой структуры. Такие подавленные эмбриогенетические системы создают возможность для приобретения новых структур, если они реактивируются на новом месте или на какой-то другой стадии развития.
Изменение местоположения индукционной системы на ранних стадиях развития может оказать глубокое воздействие на строение тела. Изменения локализации тех или иных структур сопровождаются их перемещением относительно других структур, уменьшением или увеличением относительных размеров и увеличением или уменьшением числа повторяющихся элементов в метамерных структурах. Мы обсудим этот последний вид изменений на примере своеобразной симметрии иглокожих. Иглокожие обладают рядом единственных в своем роде признаков, но главная загадка для всех, кто занимается их филогенезом, - это происхождение пятилучевой симметрии, характерной для всех ныне живущих и для большинства вымерших классов иглокожих.
Стратман (Strathmann) изучал ограничения морфологического разнообразия, налагаемые на иглокожих амбулакральной системой. В этой системе имеются такие структуры, как амбулакральные ножки или брахиолы, участвующие в дыхании, передвижении и питании. В процессе эволюции иглокожих происходило увеличение их размеров и они оказались перед физической дилеммой: для того чтобы с увеличением размеров форма тела оставалась неизменной, длина и площадь амбулакральной системы по отношению к объему тела должны уменьшаться. Компенсаторное увеличение размеров амбулакральной системы достигается в разных группах иглокожих путем увеличения длины или ширины амбулакров, а также путем их ветвления или скручивания. К этому можно было бы добавить также изменение симметрии, однако, за исключением изменений, произошедших в ранний период истории этого типа, тип симметрии у иглокожих оставался постоянным; это позволяет считать, что для большинства классов иглокожих пятилучевой тип строения оказался в селективном отношении оптимальным. Двусторонняя симметрия, наблюдаемая у личинок до метаморфоза, рассматривается как указание на то, что предок иглокожих был двусторонне симметричным и что пятилучевая симметрия наложилась на эту изначальную симметрию (обзор соответствующих данных см. Hyman). Юбагс (Ubaghs) на основе палеонтологических данных по самым древним иглокожим высказал мнение, что наиболее примитивные и древние иглокожие обладали, вероятно, двусторонней симметрией или даже были асимметричными. Хауг (Haugh) и Белл (Bell), опираясь на результаты своих недавних исследований фоссилизированных внутренних органов вымерших иглокожих, утверждают не только, что пятилучевая симметрия не является основной для этого типа, но также что у некоторых вымерших классов отсутствовала система наполненных водой сосудов. Пятилучевая симметрия могла возникнуть у одной из древних форм, обладавшей небольшим числом амбулакров. Существуют вероятные кандидаты на роль таких форм. Геликоплакоидеи с их единственным раздвоенным амбулакром и паракриноидеи с парой амбулакров представляют линии, не обладавшие пяти лучевой симметрией. Белл высказал мнение, что трехлучевые стадии онтогенеза ордовикских эдриоастероидей, которые во взрослом состоянии обладают пятилучевой симметрией, возможно, представляют собой рекапитуляцию более примитивной симметрии. Быть может, загадочное трехлучевое эдиакарское животное Tribrachidium было таким предковым иглокожим. Как мы сейчас покажем, генетика развития, по крайней мере если судить по современным иглокожим, допускает возможность, что подобные эволюционные изменения происходили быстро и «легко».
Тип симметрии, т.е. число амбулакров, имеющихся у взрослого иглокожего, определяется процессом «подсчета», происходящим на ранних стадиях личиночного развития. Этот процесс иллюстрирует рис. 5-12, на котором изображено развитие гидроцеля у обобщенного зародыша иглокожих. Гидроцель и, в сущности, вся амбулакральная система развиваются из левого целомического мешка, который формируется вблизи ротового отверстия личинки. Соответствующий правый целомический мешок дегенерирует. По мере продолжения роста гидроцеля он образует пять выростов. В результате взаимодействия с лежащим над ними эпидермисом эти выросты индуцируют рост пяти радиальных амбулакров. Если, как это сделал Чихак (Czihak), левый целом 3-4-дневной личинки морского ежа разрушить локализованным облучением очень узким пучком УФ-света, то образования гидроцеля и зачатков амбулакров не происходит. У личинок, лишенных гидроцеля, амбулакры не развиваются. Чихак установил также, что в тех немногих случаях, когда гидроцели формировались как из левого, так и из правого целомических мешков, возникали двойные зародыши с двумя амбулакральными системами. Таким образом, детерминация типа симметрии зависит от регуляции меристического признака, проявляющегося у личинки на ранних стадиях развития зачатка дефинитивного органа.
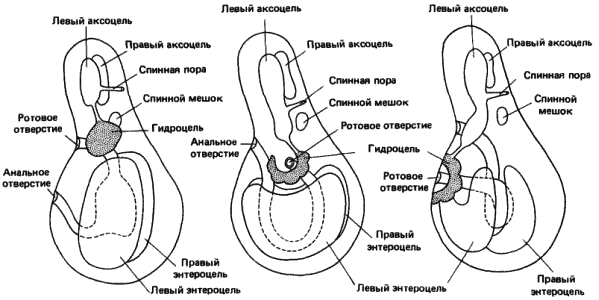
Рис. 5-12. Развитие гидроцеля (выделен пунктиром) у схематизированной личинки иглокожих, сопровождающееся образованием пяти слепых выростов (Ubaghs, 1967).
Генетический анализ механизма, лежащего в основе регуляции числа амбулакральных зачатков при развитии амбулакральной системы, затрудняется тем, что иглокожие - неподходящий объект для генетических экспериментов. Тем не менее Хайнгарднер (Hinegardner) сумел создать методы для выращивания морских ежей в лаборатории и получил несколько мутантов. Наиболее загадочным из них был мутант с измененной симметрией, которого Хайнгарднер назвал square - квадрат (рис. 5-13). У таких мутантов симметрия четырех-, а не пятилучевая, так что у них имеются 4 амбулакра, а аристотелев фонарь содержит четыре элемента. Как показали приведенные Хайнгарднером генетические данные, этот меристический признак контролируется более чем одним геном. Кроме того, данная мутация не просто обусловливает четырехлучевую симметрию как таковую; скорее, она, по-видимому, приводит к утрате контроля над симметрией. Чаще всего мутанты обладают четырехлучевой симметрией, однако от одного и того же скрещивания иногда возникают потомки с двух-, трех-, четырех-, пяти- и шестилучевой симметрией. Из всех таких отклоняющихся форм жизнеспособны только четырехлучевые, потому что у взрослых особей с нарушениями симметрии других типов аристотелев фонарь функционировать не может. Хук (Houke) и Хайнгарднер изучали морфогенетическую основу действия мутации square у живых зародышей. В то время как у нормальных личинок растущий гидроцель образует ожидаемые пять слепых выростов, у мутантных особей этого не происходит. Обычно у них появляются только четыре выроста. После метаморфоза из таких аномальных личинок получаются взрослые особи с соответствующей аномальной симметрией. Морские ежи в процессе своей эволюции, очевидно, не использовали мутационные изменения этой генетической системы в отличие от морских звезд, которые их использовали. У ряда видов морских звезд число рук и связанных с ними амбулакров увеличилось. По крайней мере некоторые из таких случаев обусловлены изменением в процессе «подсчета» этих зачатков на ранних стадиях развития. Хорошим примером служит развитие обычной для западного побережья шестилучевой морской звезды Leptasterias hexaetis. На рис. 5-14 можно видеть, как у личинки гидроцель образует пять выростов, а вскоре после этого - еще и шестой. Это приводит к образованию у ювенильной морской звезды шести рук. Подобный же тип развития (с соответствующими арифметическими поправками) происходит у девятилучевой морской звезды Solaster endica, которую описал Джемилл (Gemmill).
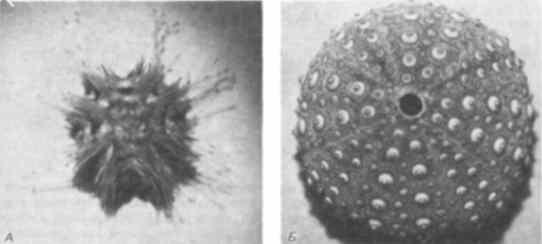
Рис. 5-13. Мутантные особи морских ежей, обладающие четырехлучевой, а не пятилучевой симметрией. А. Выращенный в лаборатории экземпляр Lytechinus pictus с четырьмя радиальными амбулакральными каналами, т.е. с фенотипом, соответствующим мутации square. Б. Найденный в природе экземпляр Strongylocentrotus franciscanus с четырехлучевой симметрией (С фотографий, любезно предоставленных R.T. Hinegardner).
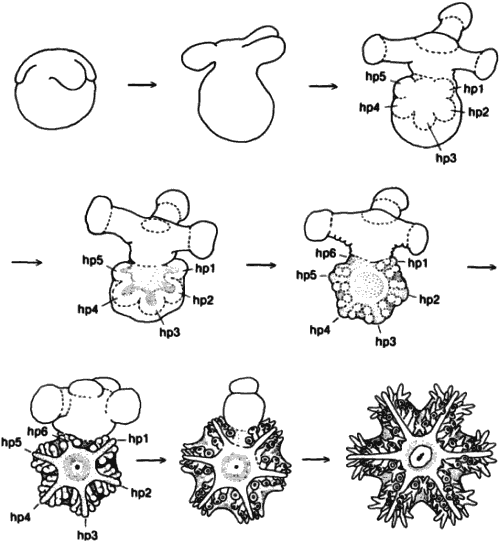
Рис. 5-14. Развитие шестилучевой морской звезды Leptasterias hexaetis; hpl-hp6-выросты гидроцеля; обратите внимание, что hp6 появляется позднее, чем hpl-hp5 (Osterud, 1918).
Генетическая основа симметрии у вымерших иглокожих, по-видимому, была сходна с генетикой морских ежей, и ископаемые мутантные бластоидеи с тремя, четырьмя и шестью амбулакрами встречаются довольно часто (рис. 5-15). Результаты исследований Ваннера (Wanner) по пермским бластоидеям и Макурда (Macurda) - по бластоидее Pentremites свидетельствуют о довольно высокой частоте мутантов с измененной симметрией (1 на 200) в популяциях этих форм. Белл описывает эдриоастероидеи с четырьмя, шестью и даже девятью амбулакрами. Иглокожие, принадлежащие к этим классам, по типу питания относились к фильтраторам, а поэтому число подвижных частей аристотелева фонаря не имело для них такого большого значения, как для морских ежей, мутантов которых изучал Хайнгарднер; это делало возможным выживание особей с резко отклоняющимися типами симметрии.
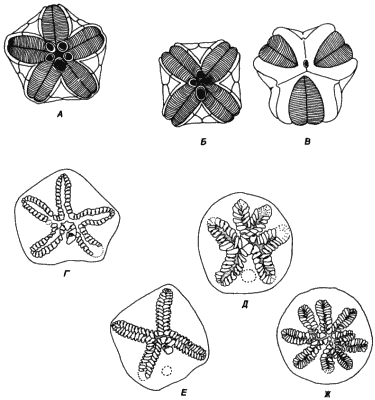
Рис. 5-15. Мутации, затрагивающие симметрию у представителей вымерших классов иглокожих. А. Нормальная пятилучевая бластоидея. Б и В. Бластоидеи, отклоняющиеся по типу симметрии. Г. Нормальная пятилучевая эдриоастероидея. Д-Ж. Эдриоастероидеи с четырьмя, шестью и девятью амбулакрами. (Бластоидеи зарисованы с экземпляров, предоставленных J.A. Waters, и перерисованы из книги Н. Н. Beaver, Morphology; см: Treatise on Invertebrate Paleontology, Part S., Echinodermata I, vol.11., 1967, R.C.Moore, ed. С любезного разрешения Американского геологического общества и Университета шт. Канзас. Эдриоастероидеи перерисованы из работы Bell, 1976a).
Непосредственное изучение генетической основы прерывистых изменений числа амбулакров у иглокожих затруднительно, но Сьюэлл Райт (Wright) провел очень тонкий анализ аналогичной ситуации - изменения числа пальцев, развивающихся у морских свинок. На передних лапах морской свинки обычно недостает первого пальца, а на задних - первого и пятого. Но иногда у морских свинок развивается 5-й палец, и путем отбора можно создать инбредные линии, у которых всегда имеется полностью сформированный 5-й палец со всеми костями, мышцами и когтем. Райт изучил несколько инбредных линий, четко различавшихся по частоте развития мизинца. Линия D, которую первоначально вывел Касл (Castle) от четырехпалого варианта из природной популяции, примерно за 20 лет до исследований Райта постоянно давала только четырехпалых потомков, линия 2 - только трехпалых. Райт проанализировал также две другие инбредные линии, 13 и 35, которые, как мы увидим, значительно отличались одна от другой и от линии 2 по генетическому составу. Для линии 13 была характерна трехпалость, как и для линии 2, тогда как популяция линии 35 примерно на 40% состояла из четырехпалых особей. Райт скрещивал линию D со всеми этими линиями. Признаки потомков F1 от каждого скрещивания представлены в табл. 5-1. От скрещивания между линиями 2 и D были получены только трехпалые потомки, скрещивание между линиями 13 и D дало значительный процент особей со слабо развитым 4-м пальцем, тогда как большая часть потомков от скрещивания между линиями 35 и D была четырехпалой.
Таблица 5-1. Число пальцев на задних лапах у морских свинок, полученных при скрещиваниях различных инбредных линий (по данным Wright, 1934)
Скрещиваемые линии Доля особей с разным числом пальцев в F1 и F2, % 3 пальца 4-й палец развит слабо 4 пальца F1 2 x D 100 0 0 13 x D 69 31 0 35 x D 6 13 81 F2 2 x D 77 16 7 13 x D 48 19 33 35 x D 25 20 55Скрещивания трехпалых особей F1 друг с другом давали трех- и четырехпалых потомков. В поколении F2 от скрещивания между собой гибридов 2 x D происходило расщепление в соответствии с менделевским отношением 3:1; это позволяет считать, что данный признак контролируется парой аллелей одного гена. Однако Райт провел ряд возвратных скрещиваний особей F2 с линией D, которые показали, что это простое объяснение не может быть верным, и что на самом деле более вероятно, что линии 2 и D различаются по целому набору аллелей, относящихся к четырем генам. Линии 13 и 35 отличаются от линии D по меньшему числу аллелей. Заключения Райта схематически изображены на рис. 5-16, где представлены графики зависимости распределения фенотипов от генотипа. Ввиду того что число пальцев - дискретный признак, существует некое пороговое число необходимых аллелей: для развития четырехпалой лапы требуется наличие не менее чем пяти аллелей полидактилии.
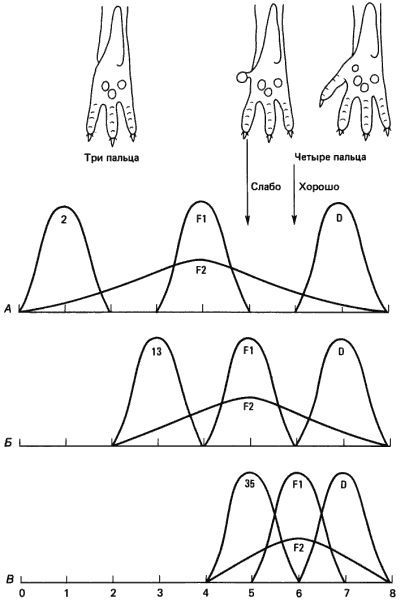
Рис. 5-16. Генетическая регуляция числа пальцев на задних лапах у морской свинки. Вверху слева - нормальная трехпалая задняя лапа морской свинки, справа - лапа с хорошо развитым 4-м пальцем, в центре - лапа со слабо развитым 4-м пальцем. Кривые иллюстрируют распределение фенотипов в зависимости от числа аллелей полидактилии в потомстве от разных скрещиваний между тремя инбредными линиями. Стрелками показаны генетические пороги между трехпалой лапой, лапой со слабо развитым и лапой с хорошо развитым четвертым пальцем. А. Скрещивание между линиями D (четырехпалая) и 2 (трехпалая); эти две линии сильно различаются по генетической конституции, и у потомков F1 число генов полидактилии недостаточно для развития у них четырех пальцев. Б. Скрещивание между линиями D (четырехпалая) и 13 (трехпалая); линия 13 содержит больше генов полидактилии, чем линия 2, а поэтому у потомков F1 образуется лапа с недоразвитым четвертым пальцем. В. Скрещивание между линиями D (четырехпалая) и 35 (дающей как трех-, так и четырехпалых потомков) (S. Wright, 1934a, b).
Рассматриваемый прерывистый признак, регулируемый таким образом у морских свинок, возникает на поздних стадиях развития и не оказывает сколько-нибудь существенного влияния на строение организма в целом. Однако такого рода генетический контроль, действующий на ранних стадиях развития, как у иглокожих, оказывает глубокое воздействие на строение тела и, возможно, сыграл решающую роль в макроэволюции классов иглокожих, обладающих пятилучевой симметрией. «Перспективные монстры» не обязательно должны быть результатом внезапных и всеобъемлющих перестроек генетической программы онтогенеза. Небольшое генетическое изменение может привести к такому изменению развития, которое допускает коренное изменение адаптации: маленьким ключом можно отпереть большую дверь.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
4.1. Типы взаимодействия
4.1. Типы взаимодействия Выделяют следующие типы взаимодействия между популяциями: «1) нейтрализм, при котором ассоциация двух популяций не сказывается ни на одной из них; 2) взаимное конкурентное подавление, при котором обе популяции активно подавляют друг друга; 3)
§ 1. Взаимодействия с внешней средой
§ 1. Взаимодействия с внешней средой Растение — самое большое и самое живучее существо на этой планете. Мамонтово дерево и Гигантский эвкалипт достигают высоты 120–150 м, а Долговечная сосна — возраста 4600 лет. Среди растений много крупных организмов, поэтому существуют
10. Морфология вирусов, типы взаимодействия вируса с клеткой
10. Морфология вирусов, типы взаимодействия вируса с клеткой Вирусы – микроорганизмы, составляющие царство Vira.Вирусы могут существовать в двух формах: внеклеточной (вириона) и внутриклеточной (вируса).По форме вирионы могут быть: округлыми, палочковидными, в виде
3. ИНДИВИДУАЛЬНЫЕ ОСОБЕННОСТИ ВЗАИМОДЕЙСТВИЯ СТРУКТУР МОЗГА, РЕАЛИЗУЮЩИХ ФУНКЦИИ ЭМОЦИЙ КАК ОСНОВУ ТЕМПЕРАМЕНТОВ
3. ИНДИВИДУАЛЬНЫЕ ОСОБЕННОСТИ ВЗАИМОДЕЙСТВИЯ СТРУКТУР МОЗГА, РЕАЛИЗУЮЩИХ ФУНКЦИИ ЭМОЦИЙ КАК ОСНОВУ ТЕМПЕРАМЕНТОВ В качестве методов диагностики устойчивых (типологических) индивидуальных особенностей поведения мы использовали две поведенческие модели: тест на
5.4. Макроэволюция
5.4. Макроэволюция Макроэволюция – это эволюция надвидовых таксонов. Как раздел эволюционной теории она в настоящее время не представляет собой единую и стройную систему. Скорее, это несколько теорий, в той или иной степени связанных между
Глава 5 Взаимодействия внутри зародыша
Глава 5 Взаимодействия внутри зародыша Самый акт становления, однако, не является ни абстрактным, ни мгновенным... Д. Сипфл «К вопросу о вразумительности эпохальной теории времени» Взаимодействие и интеграция В наши дни посетители музеев воспринимают выставленные в них
Глава 4. Морфогенетические поля
Глава 4. Морфогенетические поля 4.1. Морфогенетические зародыши Морфогенез не происходит в вакууме. Он может начаться только с уже организованной системы, которая служит морфогенетическим зародышем. В процессе морфогенеза новая морфическая единица более высокого уровня
4.1. Морфогенетические зародыши
4.1. Морфогенетические зародыши Морфогенез не происходит в вакууме. Он может начаться только с уже организованной системы, которая служит морфогенетическим зародышем. В процессе морфогенеза новая морфическая единица более высокого уровня возникает вокруг этого
4.3. Морфогенетические поля как вероятностные структуры
4.3. Морфогенетические поля как вероятностные структуры Орбитали электронов вокруг атомного ядра можно рассматривать как структуры в морфогенетическом поле атома. Эти орбитали могут быть описаны решениями уравнения Шредингера. Однако, согласно квантовой механике,
4.5. Морфогенетические зародыши в биологических системах
4.5. Морфогенетические зародыши в биологических системах На клеточном уровне зародыши морфогенетических трансформаций могут быть морфическими единицами низшего уровня в клетках, которые присутствуют как в начале, так и в конце процесса клеточной дифференциации.
7.2. Измененные морфогенетические зародыши
7.2. Измененные морфогенетические зародыши Морфогенетические зародыши для развития органов и тканей состоят из клеток или групп клеток с характерными структурой и организацией колебаний (разделы 4.5,6.1). Если в силу необычных условий окружающей среды или генетических
9.6. Морфогенетические поля и моторные поля
9.6. Морфогенетические поля и моторные поля Несмотря на то что поля, контролирующие изменения формы специализированных двигательных структур животных, — это фактически морфогенетические поля, они вызывают, скорее, движения, нежели изменения формы. По этой причине
МАКРОЭВОЛЮЦИЯ
МАКРОЭВОЛЮЦИЯ Жизненная стратегия пионеров рассчитана на неустойчивые условия, в которых высокая смертность сдерживает рост численности и она не достигает критических пределов, грозящих истощением природных ресурсов. Поэтому острой конкуренции не возникает. Пионеры
2.2.4. Определение способа взаимодействия хозяина с собакой
2.2.4. Определение способа взаимодействия хозяина с собакой Поскольку зачастую установлением причины девиантного поведения животного занимается врач-ветеринар общего профиля, который не обладает профессиональной психологической подготовкой, автором предлагается тест,