7.2. Измененные морфогенетические зародыши
7.2. Измененные морфогенетические зародыши
Морфогенетические зародыши для развития органов и тканей состоят из клеток или групп клеток с характерными структурой и организацией колебаний (разделы 4.5,6.1). Если в силу необычных условий окружающей среды или генетических изменений структура и организация колебаний зародыша достаточно изменились, то он перестает быть связанным с обычным для него морфогенетическим полем: тогда он либо может вообще стать неспособным действовать как зародыш, и в этом случае вся соответствующая структура не появится в организме; либо он может оказаться связанным с другим морфогенетическим полем, и тогда в этой части организма может развиться иная структура, а не та, которая обычно здесь образуется.
Было описано множество примеров такой потери целостной структуры или замены одной структуры на другую. Иногда одинаковые изменения могут быть вызваны генетическими факторами и изменениями в окружении развивающегося организма; последние в генетической литературе именуются фенокопиями.[147]
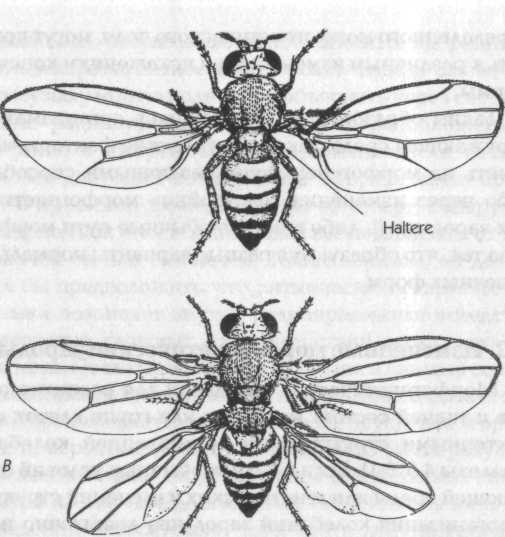
Рис. 17. Нормальная особь плодовой мушки Drosophila (А) и мутантная мушка (В), у которой третий грудной сегмент был трансформирован таким образом, что он стал похож на второй грудной сегмент. В результате мушка имеет две пары крыльев вместо одной
Эффекты такого рода были детально изучены на плодовой мушке дрозофиле. Значительное число идентифицированных мутаций приводит к трансформациям целых участков тела мушки; например, антеннопедия заменяет усики на ножки, а мутации в грудном (бито-раксовом) генном комплексе приводит к тому, что третий (задний) грудной сегмент, который в норме содержит два зачатка (halteres), развивается так, как если бы он был средним сегментом (рис. 17). При этом взрослые организмы имеют две пары крыльев на соседних сегментах.[148]
Сопоставимые явления были обнаружены у растений. Например, у гороха в норме листья включают собственно листочки у основания и усики на конце.
В некоторых листьях усики образуются напротив листьев, указывая на то, что одинаковые примордии (зачаточные листы) могут давать структуры обоих типов (рис. 18); предположительно на клетки в этих примордиях влияют некие факторы в эмбриональном листе, заставляющие эти клетки принимать структуру и организацию колебаний, характерные для морфогенетического зародыша либо усика, либо листочка. Однако в мутанте одного типа способность образовывать усики подавлена, и все примордии дают листочки; в мутанте другого типа (вследствие гена в другой хромосоме) подавлено образование листочков, и все примордии дают только усики[149] (рис. 18).
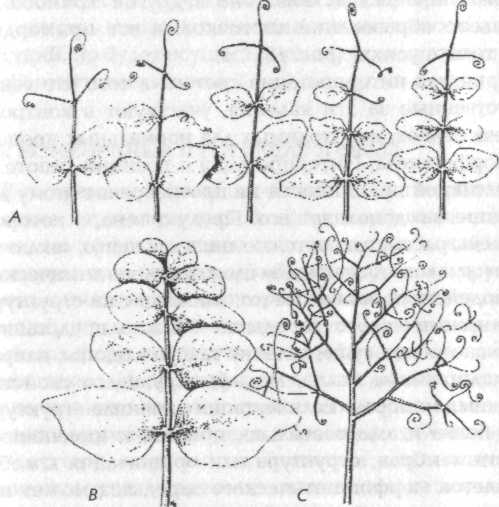
Рис. 18. А — нормальные листья гороха, имеющие как листочки, так и усики;
В — лист мутанта, у которого образуются только листочки;
С — лист мутанта, у которого образуются только усики
Принятая интерпретация состоит в том, что гены, ответственные за эти явления, участвуют в контроле синтеза белков, необходимых для нормальных процессов морфогенеза. Интерпретация с позиции гипотезы формативной причинности не противоречит этому допущению, но дополняет его. Продукт гена, о котором идет речь, рассматривается не лишь как нечто, «включающее» или «выключающее» сложную цепь химических взаимодействий, но как нечто, влияющее на структуру морфогенетического зародыша. Можно представить много способов осуществления такого влияния, например кодирование белка, модифицирующего свойства клеточных мембран. Если мутация изменила структуру этого белка и, следовательно, привела к изменениям свойств мембран, структура или организация колебаний клеток морфогенетического зародыша может измениться таким образом, что они уже не могут быть связаны с обычным для них морфогенетическим полем. Следовательно, весь путь морфогенеза будет заблокирован. Поскольку клетки, участвующие в этом пути, более не претерпевают нормальное развитие и дифференциацию, они не будут синтезировать белки, характерные для этих процессов. А если морфогенетический зародыш изменился так, что он оказался связанным с другим морфогенетическим полем с помощью морфического резонанса, то развивающиеся клетки будут синтезировать белки, соответствующие другому морфогенетическому процессу.
Таким образом, мутация, в результате которой один из путей морфогенеза оказался заблокированным или которая привела к другому пути, действительно изменяет продукт гена, косвенно контролирующий заданную схему синтеза белков, как и предполагает механистическая теория. Однако этот контроль не будет зависеть только от сложных химических взаимодействий, но будет осуществляться также посредством морфогенетических полей.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Глава 1 Зародыши и предки
Глава 1 Зародыши и предки Вероятно, мне следует пояснить, — добавил барсук, нервно опуская свои бумаги и глядя поверх них на бородавку, — что все зародыши выглядят в общем одинаково. Зародыш - это то, что вы есть прежде, чем вы родитесь на свет. И станете ли вы в будущем
Морфогенетические взаимодействия и макроэволюция
Морфогенетические взаимодействия и макроэволюция Приведенные выше примеры, относящиеся к амфибиям и птицам, иллюстрируют эволюционные последствия изменений, вызванных взаимодействиями на относительно поздних стадиях развития. Соответственно достигаемые при этом
Глава 4. Морфогенетические поля
Глава 4. Морфогенетические поля 4.1. Морфогенетические зародыши Морфогенез не происходит в вакууме. Он может начаться только с уже организованной системы, которая служит морфогенетическим зародышем. В процессе морфогенеза новая морфическая единица более высокого уровня
4.1. Морфогенетические зародыши
4.1. Морфогенетические зародыши Морфогенез не происходит в вакууме. Он может начаться только с уже организованной системы, которая служит морфогенетическим зародышем. В процессе морфогенеза новая морфическая единица более высокого уровня возникает вокруг этого
4.3. Морфогенетические поля как вероятностные структуры
4.3. Морфогенетические поля как вероятностные структуры Орбитали электронов вокруг атомного ядра можно рассматривать как структуры в морфогенетическом поле атома. Эти орбитали могут быть описаны решениями уравнения Шредингера. Однако, согласно квантовой механике,
4.5. Морфогенетические зародыши в биологических системах
4.5. Морфогенетические зародыши в биологических системах На клеточном уровне зародыши морфогенетических трансформаций могут быть морфическими единицами низшего уровня в клетках, которые присутствуют как в начале, так и в конце процесса клеточной дифференциации.
7.3. Измененные пути морфогенеза
7.3. Измененные пути морфогенеза В то время как факторы, влияющие на морфогенетические зародыши, производят в морфогенезе качественные эффекты, такие как отсутствие какой-либо структуры или замена одной структуры на другую, многие генетические факторы или воздействия
9.6. Морфогенетические поля и моторные поля
9.6. Морфогенетические поля и моторные поля Несмотря на то что поля, контролирующие изменения формы специализированных двигательных структур животных, — это фактически морфогенетические поля, они вызывают, скорее, движения, нежели изменения формы. По этой причине