9.2. Движения растений
9.2. Движения растений
Обычно растения движутся путем роста.[181] Этот факт легче осознать, когда видишь их в ускоренной киносъемке: ростки вытягиваются и изгибаются к свету; разветвления корней устремляются вниз, в почву, а верхушки усиков и ползучих стеблей выбрасывают в воздух широкие спирали, пока не соприкоснутся с твердой опорой и не обовьются вокруг нее.[182]
Рост и развитие растений происходят под контролем их морфогенетических полей, которые сообщают растениям их характерные формы. Но ориентация этого роста в значительной степени определяется направленными стимулами силы тяжести и света. Факторы окружения влияют также на тип развития: например, в тусклом свете растения становятся белесыми, их побеги растут сравнительно быстро и становятся длинными и тонкими, пока не достигнут более яркого света.
Гравитация «чувствуется» благодаря ее действию на зерна крахмала, которые скатываются вниз и накапливаются в нижних частях клеток.[183] Направление, откуда приходит свет, обнаруживается путем дифференциального поглощения лучистой энергии на освещенной и теневой сторонах органов желтым пигментом каротеноидом.[184] Чувство «осязания», с помощью которого ползучие стебли и усики находят твердые опоры, может объясняться выделением простого химического вещества, этилена, с поверхности клеток при их механической стимуляции.[185] Изменение роста с ускоренного при недостатке света на нормальный зависит от поглощения света голубым белковым пигментом, называемым фитохромом.[186]
Ответы на эти стимулы включают сложные физико-химические изменения в клетках и тканях, а в некоторых случаях зависят от различий в распределении гормонов, таких как ауксин. Однако эти реакции не могут быть объяснены с помощью одних только физико-химических изменений, но могут быть поняты лишь с привлечением общих морфогенетических полей растений. Например, благодаря присущей им полярности растения производят на одном конце стебли, на другом — корни. Направленный стимул гравитации руководит этим поляризованным развитием так, что стебли растут вверх, а корни вниз. Действие гравитационного поля на зерна крахмала в клетках и соответствующие изменения в распределении гормонов действительно являются причиной этих ориентированных движений роста, но не могут сами по себе объяснить существование полярности; не объясняют они и тот факт, что главные стебли и корни реагируют диаметрально противоположным образом, а также различные особенности роста деревьев, трав, вьющихся и ползучих растений и особенности разветвления стебля и корневой системы различных видов. Все эти признаки зависят от морфогенетических полей.
Несмотря на то что большая часть движений растений происходит только в молодых растущих органах, некоторые структуры сохраняют способность двигаться даже тогда, когда они вполне созрели, например, цветы, которые открываются и закрываются каждый день, и листья, складывающиеся ночью. На эти движения влияют интенсивность света и другие факторы среды; они находятся также под контролем «физиологических часов» и продолжают происходить приблизительно раз в сутки, даже если растения помещаются в неизменяемые условия.[187] Листья или лепестки открываются, потому что специализированные клетки в «шарнирной» (hinge) области у их основания разбухают; и они закрываются, когда эти клетки теряют воду вследствие изменения проницаемости их мембран по отношению к неорганическим ионам.[188] Восстановление тургорного давления является активным процессом, требующим затрат энергии и сравнимым с ростом.
Помимо «сонных» движений листья некоторых растений движутся в течение дня в ответ на изменение положения солнца. Например, у голубиного гороха, Cajanus cajan, листочки, на которые падает солнце, ориентированы приблизительно параллельно солнечным лучам, что минимизирует площадь поверхности, подверженной интенсивному тропическому излучению. Но листья в тени ориентируются под прямыми углами к падающим лучам, тем самым улавливая максимальное количество света. Эти реакции зависят от направления и интенсивности света, падающего на специализированные узлы в листьях (листовые подушечки). В течение дня листья и листочки непрерывно приспосабливают свое положение к движению солнца по небу. Ночью они принимают свое вертикальное «спящее» положение: листовые подушечки чувствительны к гравитации так же, как и к свету.
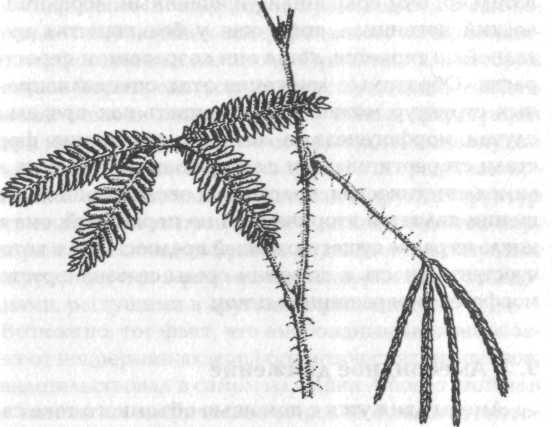
Рис. 23. Листья чувствительного растения, Mimosa pudica. Слева — не стимулированные; справа — стимулированные
У «чувствительного» растения, Mimosa pudica, листочки закрываются и листья смотрят вниз ночью, как и у многих других бобовых растений. Но эти движения быстро происходят также и в дневное время в ответ на механический стимул {рис. 23). Стимул вызывает распространение вниз по листу волны электрической деполяризации, подобной нервному импульсу; если стимул достаточно сильный, импульс распространяется на другие листья, которые также свертываются.[189] Подобно этому, у растения венерина мухоловка, Dionaea muscipula, механическая стимуляция чувствительных волосков на поверхности листа вызывает движение электрического импульса к разбухшим «шарнирным» клеткам, которые быстро теряют воду; в результате лист захлопывается как ловушка вокруг несчастных насекомых, которые затем перевариваются.[190]
Эти движения листьев и листочков в ответ на действие света, силы тяжести и механическую стимуляцию возможны потому, что специализированные клетки способны терять воду, а затем вырастать снова; следовательно, они сохранили упрощенный морфогенетический потенциал, тогда как у большинства других тканей он теряется, когда они созревают и перестают расти. Обратимые движения этих специализированных структур можно рассматривать как предельные случаи морфогенеза, в которых изменения формы стали стереотипными и повторяющимися. Но их квазимеханистическая простота в эволюционном отношении является вторичной, а не первичной: она возникла из ранее существовавшей предпосылки, в которой чувствительность к стимулам среды связана с ростом и морфогенезом растения в целом.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
КЛАССИФИКАЦИЯ РАСТЕНИЙ ОСНОВНЫЕ СИСТЕМАТИЧЕСКИЕ ГРУППЫ. ГЛАВНЫЕ ПРИЗНАКИ ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ. ДВУДОЛЬНЫЕ И ОДНОДОЛЬНЫЕ
КЛАССИФИКАЦИЯ РАСТЕНИЙ ОСНОВНЫЕ СИСТЕМАТИЧЕСКИЕ ГРУППЫ. ГЛАВНЫЕ ПРИЗНАКИ ПОКРЫТОСЕМЕННЫХ РАСТЕНИЙ. ДВУДОЛЬНЫЕ И ОДНОДОЛЬНЫЕ 1. Систематика – это наука, изучающаяA. Происхождение растительного мираБ. Строение живых организмовB. Приспособление особей к окружающей
РАЗВИТИЕ РАСТИТЕЛЬНОГО МИРА ПРОИСХОЖДЕНИЕ РАСТЕНИЙ. ОХРАНА РАСТЕНИЙ
РАЗВИТИЕ РАСТИТЕЛЬНОГО МИРА ПРОИСХОЖДЕНИЕ РАСТЕНИЙ. ОХРАНА РАСТЕНИЙ 1. Палеонтология – это наука, изучающая:A. Строение растительных организмовБ. Геологическое строение ЗемлиB. Условия средыГ. Вымершие организмы и смену их во времени2. Первые живые организмы появились:A.
В чем состоит главное отличие движения Венеры и Урана от движения остальных планет?
В чем состоит главное отличие движения Венеры и Урана от движения остальных планет? Все планеты обращаются вокруг Солнца в одном направлении – в том же, в котором вращается вокруг своей оси Солнце. В этом же направлении вращаются почти все планеты и вокруг собственных
ДВИЖЕНИЯ
ДВИЖЕНИЯ Только при наличии хорошего ходового аппарата и выносливости можно использовать физиологические способности собаки для той или иной цели. Движение собаки — ее манеру и легкость движения — очень часто недооценивают при экспертизе, предпочитая судить о
Движения
Движения Когда щенкам два — три дня от роду можно заметить, что они уже достаточно сильные, живые и могут ползать, продвигаясь вперед неуверенными толчками. Сначала они ползают медленно, качаясь и бросая голову из стороны в сторону, как бы стараясь удержать
Движения
Движения Когда щенкам два — три дня от роду можно заметить, что они уже достаточно сильные, живые и могут ползать, продвигаясь вперед неуверенными толчками. Сначала они ползают медленно, качаясь и бросая голову из стороны в сторону, как бы стараясь удержать
Инстинктивные движения
Инстинктивные движения Стабильные, жесткие инстинктивные движения появляются в раннем постнатальном онтогенезе в настолько «готовом» виде, что долгое время считалось, что они совершенно не развиваются и не нуждаются в индивидуальном упражнении. На самом деле, как мы
Движения простейших
Движения простейших Движения простейших отличаются большим разнообразием, причем в этом типе простейших животных встречаются способы локомоции, которые совершенно отсутствуют у многоклеточных животных. Это своеобразный способ передвижения амеб при помощи
Движения
Движения Кольчатые черви обитают в морях и пресноводных водоемах, но некоторые ведут и наземный образ жизни, передвигаясь ползком по субстрату или роясь в рыхлом грунте. Морские черви отчасти пассивно носятся течениями воды как составная часть планктона, но основная
Движения
Движения Огромное разнообразие «экологических ниш», занимаемых насекомыми и другими членистоногими, обусловило, естественно, развитие весьма различных форм приспособления в области строения и поведения этих животных. Наиболее существенным и характерным для эволюции
Движения глаз во сне?
Движения глаз во сне? То ли дело быстрые движения глаз! Нет сомнения, что они означают «смотрение» снов. Как это доказать? В ходе экспериментов Клейтман и Демент научились по рассказам о сновидениях, предшествующих пробуждению, угадывать, какие движения глаз можно ожидать