Время и генетическая дивергенция
Время и генетическая дивергенция
Скорости и способы видообразования очень удобно рассмотреть на примере большой группы видов сем. Drosophilidae, эндемичных для Гавайских островов, которую так отлично изучил Карсон (Н. Carson) со своими сотрудниками и студентами. Наш выбор диктуется несколькими исключительными особенностями как Гавайского архипелага, так и обитающих на нем дрозофил. Эти острова хорошо изолированы от материка и поэтому, так же как и дарвиновские Галапагосы, могут служить природной лабораторией для исследования эволюции. Кроме того, геологический возраст разных островов, составляющих Гавайский архипелаг, хорошо установлен калий-аргоновым и палеомагнитным методами. Если двигаться по современной карте архипелага (рис. 3-7) с севера на юг, то острова становятся все моложе. Это обусловлено перемещением тихоокеанской литосферной плиты над «горячей точкой» под поверхностью Земли.
По мере миграции плиты (на северо-запад, со скоростью примерно 9 см/год) над этим местом возникал ряд вулканов. Так, остров Кауаи появился в плиоцене, примерно 5 млн. лет назад, а Оаху - 3 млн. лет назад. Три небольших острова - Молокаи, Мауи и Ланаи, - первоначально составлявшие одно целое, образовались 1,5 млн. лет назад, а позднее разделились. Наконец, самый большой и самый молодой остров Гавайи все еще находится над «горячей точкой», о чем свидетельствует продолжающаяся вулканическая активность. Самые древние части этого самого молодого острова сформировались в плейстоцене, несколько менее 1 млн. лет назад. Последовательное возникновение этих островов в сочетании с их изолированностью как от азиатского, так и от североамериканского материков создали такую ситуацию, которая позволяет проследить весь процесс заселения новой территории и новых ниш, изучая ныне существующие виды организмов. Дальнейшее преимущество связано с тем, что Гавайские острова находятся в тропиках, и поэтому их биота весьма разнообразна. Это ясно видно на примере сем. Drosophilidae, которое нам предстоит рассмотреть. По оценкам Карсона (Carson) и Канеширо (Kaneshiro), на относительно небольшой территории названных выше шести островов встречается более 500 эндемичных видов этих мух. Отсюда следует, что то событие, которое мы собираемся изучать, т.е. видообразование, происходило в этой изолированной, охарактеризованной во времени, среде с достаточной регулярностью.
Другой и столь же важный аспект рассматриваемого исследования - это организм, служивший его объектом. Дрозофилы не только позволяют подвергнуть анализу большое число видов; но в силу их генетических особенностей получить информацию об их филогенетических взаимоотношениях легче, чем для большинства других организмов. В этом смысле особенно удобны их политенные хромосомы. Почти во всех личиночных тканях этих двукрылых имеются клетки, в которых происходит репликация ДНК, не сопровождающаяся клеточным делением.
Реплицировавшиеся интерфазные хромосомы увеличиваются до такой степени, что их можно исследовать под световым микроскопом. При этом видно, что они состоят из ряда интенсивно окрашивающихся полос-дисков, разделенных более светлыми междисками. Каждая хромосома каждого данного вида отличается от других собственным очень характерным расположением широких и узких полос по всей своей длине. Сравнивая характер полосатости (бендинг) у разных видов, можно выявить перераспределения генетического материала в их хромосомах. Это упрощается благодаря существованию у них, как и у других двукрылых, соматической конъюгации гомологичных хромосом. Кроме того, в тех случаях, когда удается получить потомство от какого-либо межвидового скрещивания, можно исследовать политенные хромосомы потомков и выявить наличие у них генетических перестроек. Этот метод можно дополнить изучением метафазных хромосом других более обычных клеток рассматриваемых видов.
Карсон и Клейтон (F. Clayton) в течение нескольких лет изучали политенные и метафазные хромосомы видов Drosophilidae, эндемичных для Гавайских островов. Они обнаружили, что преобладающее большинство изученных до сих пор видов имеют одинаковый основной метафазный кариотип - пять палочковидных хромосом и одну точечную, а диплоидное число хромосом равно у них 12. Результаты изучения политенных хромосом, позволяющих гораздо детальнее рассмотреть морфологию хромосом, не подтверждают этот кажущийся консерватизм, так как выявляют большое число хромосомных инверсий. Интересно, что эти инверсии представляют собой не просто случайные перестройки хромосом, возникшие у разных видов, а образуют определенные группы и подгруппы; т.е. если произвольно принять последовательность хромосомных дисков у какого-либо вида за стандартную, то другие виды можно сравнивать с этим стандартом, выясняя, по скольким инверсиям они различаются. По мере сопоставления все большего числа «нестандартных» видов со стандартом в наблюдаемых различиях начинает выявляться определенная упорядоченность и возникает возможность построить некую филогению, допустив, например, что если вид А отличается от стандарта группой из пяти или шести инверсий, а вид В - этими же пятью-шестью и вдобавок еще тремя, то представляется вероятным, что вид В находится в более отдаленном родстве со стандартом, чем вид А. Именно таким способом Карсон сумел построить филогенетическое древо для более чем 100 видов гавайских Drosophilidae.
Небольшой участок этого основанного на хромосомных различиях древа представлен на рис. 3-7; он соответствует так называемой группе planitibia этих дрозофил. На кладограмме показаны хромосомные взаимосвязи для 8 из 17 описанных видов этой группы. Если сопоставить приведенную кладограмму с распределением этих 8 видов по разным островам, то выявляется интересная закономерность, позволяющая ввести в созданную схему возможную временную компоненту. Drosophila picticornis встречается только на Кауаи - самом древнем из всех островов, и, судя по структуре его хромосом, этот вид связан с D. attigua и D. primaeva более близким родством, чем другие члены группы planitibia. Считается, что эти два вида относятся к числу самых давних обитателей Гавайских островов и наиболее близки к материковому виду, который, согласно предположениям, первым заселил эти острова и положил начало всей линии гавайских представителей Drosophilidae. На острове Оаху обитают два вида - D. substenoptera и D. hemipeza. Первый из них ближе к D. picticornis и, вероятно, произошел от какого-то сходного с D. picticornis предка, который колонизировал остров Оаху вскоре после его образования, что произошло спустя примерно 2 млн. лет после возникновения острова Кауаи. Судя по морфологии его хромосом, вид D. hemipeza, по-видимому, не связан прямым родством ни с D. picticornis, ни с D. substenoptera, а происходит от какого-то предка, который был сходен с D. neopicta и вторично мигрировал с острова Молокаи обратно на Оаху. Эта гипотеза совместима с наличием у D. hemipeza группы инверсий, сходных с инверсиями, имеющимися у D. neopicta, а также двух других инверсий в Х-хромосоме, одна из которых встречается только у D. hemipeza. Прямое выведение D. hemipeza от D. substenoptera нарушило бы принцип экономии мышления, принятый при построении филогенетического древа. Кроме того, этот аспект филогении хромосом подтверждается биохимическими данными, полученными при анализе полиморфизма по ферментам у этих видов, которые мы приведем чуть ниже.
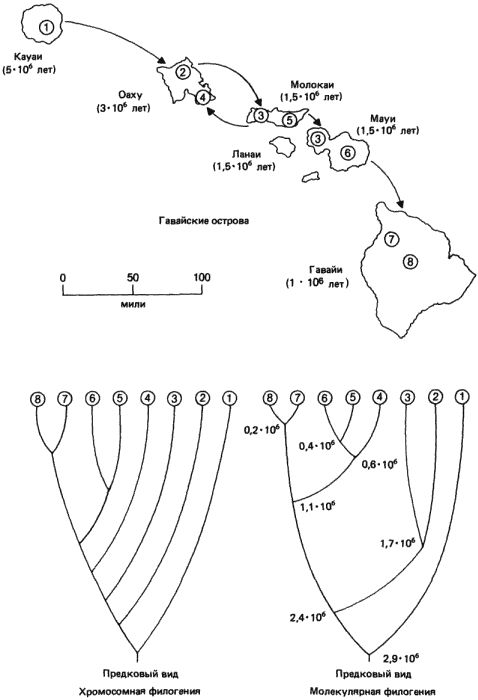
Рис. 3-7. Карта основных островов Гавайского архипелага. Под названием каждого острова в скобках указано время его возникновения (судя по возрасту слагающих его вулканических пород). Стрелками между островами показано направление миграций и случаи основания новых популяций, предположительно имевшие место в процессе эволюции гавайских Drosophilidae. Под картой помещены две дендрограммы, изображающие филогению части группы plantibia. Левая дендрограмма основана на данных об инверсиях, наблюдаемых в политенных хромосомах этих видов. Правая дендрограмма построена по данным вычислений, в основе которых лежит сходство электрофоретической подвижности ферментов, кодируемых 10 разными локусами (сами коэффициенты сходства представлены в табл. 3-2). Значения времени, указанные в каждой точке дивергенции, основаны на определяемой по молекулярным часам скорости замещения аминокислот в исследованных ферментах. Для установления средней скорости этого процесса использовали время возникновения разных островов. Филогении восьми видов, созданные этими двумя независимыми способами, почти идентичны; обе они соответствуют концепции, согласно которой мухи мигрировали с острова на остров, основывая в период заселения островов новые виды. Цифрами в кружках и на дендрограммах обозначены локализация и филогенетические взаимоотношения следующих видов: 1) D. picticornis; 2) D. substenoptera; 3) D. neopicta; 4) D. hemipeza; 5) D. differens; 6) D. planitibia; 7) D. silvestris; 8) D. heteroneura (Johnson et al., 1975).
На острове Молокаи обитает также D. differens, а на соседнем острове Мауи, когда-то соединявшемся с Молокаи, - вид D. planitibia, давший название всей группе. Оба этих вида идентичны по структуре политенных хромосом, и тем не менее они изолированы (как репродуктивно, так и географически). Поскольку D. neopicta обитает на обоих островах, представляется вероятным, что либо D. differens, либо D. planitibia произошли в процессе видообразования от D. neopicta. Какой из этих двух видов, идентичных по своим хромосомам, дал начало другому - менее ясно; нет данных, которые подтверждали бы, что D. differens дал начало D. planitibia, мигрировав с острова Молокаи на Мауи и колонизировав последний, или же что произошло обратное.
Последняя пара изучавшихся видов, D. silvestris и D. heteroneura, встречаются только на самом большом и самом молодом острове Гавайи. Судя по структуре хромосом этих двух видов, которая у них идентична (так же, как у D. differens и D. planitibia), они могли возникнуть в результате колонизации Гавайи каким-то предком, сходным с D. planitibia. Два вида, обитающие на большом острове, существуют почти симпатрически, и тем не менее в природных условиях они редко скрещиваются между собой. В лаборатории, однако, их удается скрестить, получая жизнеспособное и плодовитое потомство. Это, казалось бы, бросает вызов нашему первоначальному определению вида. И все же морфологические различия между этими видами, которые будут описаны в дальнейшем, а также их изоляцию в природе следует рассматривать как указание на то, что их разделение произошло лишь совсем недавно.
Факт разделения этих двух симпатрических видов в природе выносит на поверхность главную, хотя и не единственную, причину изоляции среди этих видов. Главный механизм изоляции - презиготический, и он связан с рядом аспектов полового и репродуктивного поведения. У самцов многих видов гавайских Drosophilidae наблюдается ярко выраженное территориальное поведение, и они защищают свою территорию от вторжения чужих особей. Они заманивают самок на эти «тока», или репродуктивные территории, после чего происходит очень сложная церемония ухаживания и спаривания. Изолирующие факторы, а поэтому и процессы видообразования в этой группе организмов по большей части обусловлены изменениями в брачном поведении в сочетании с морфологическими изменениями, связанными с полом.
У многих идентифицированных видов дрозофилы, а в особенности у членов группы planitibia, была исследована степень изменчивости ферментных белков и ее типы. У восьми, предполагаемая филогения которых представлена на рис. 3-7, определяли и сравнивали электрофоретическую подвижность ряда их ферментов. На основании таких данных по 10 ферментам Карсон и его сотрудники вычислили коэффициент сходства для всех попарных сравнений изучавшихся 8 видов. Полученные значения приведены в табл. 3-2.
Таблица 3-2. Коэффициенты сходства, полученные при попарных сравнениях восьми гавайских видов Drosophila
(Johnson et al., 1975)
sil het pla dif hem neo sub pic silvestris 1,00 0,96 0,74 0,71 0,56 0,39 0,30 0,22 heteroneura 1,00 0,76 0,72 0,58 0,41 0,30 0,23 planitibia 1,00 0,85 0,78 0,49 0,40 0,39 differens 1,00 0,74 0,50 0,40 0,36 hemipeza 1,00 0,40 0,40 0,48 neopicta 1,00 0,59 0,39 substenoptera 1,00 0,29 picticornis 1,00Значение 1,00, полученное при сравнении D. silvestris и D. silvestris, означает генетическую идентичность по всем ферментам, тогда как значение 0,00 означало бы полное отсутствие какого бы то ни было сходства. Как показывает эта таблица, два симпатрических вида с самого большого и самого молодого острова Гавайи очень близки друг другу. Для того чтобы читатель имел эталон для сравнения, приведем данные, полученные Добржанским и его сотрудниками, изучавшими комплекс D. willistoni из Центральной и Южной Америки. По этой группе были проделаны аналогичные исследования, сравнения и вычисления. Среди этих мух Добржанский с сотрудниками выделили группы, которые они считают популяциями, подвидами, зарождающимися видами, видами-двойниками и морфологически различимыми видами (в порядке убывания таксономического сходства). Величины S, или коэффициенты сходства, для этих пяти категорий составили 0,97; 0,79; 0,79; 0,56 и 0,35 соответственно. Сравнивая эти значения с приведенными в табл. 3-2, мы убеждаемся, что D. silvestris и D. heteroneura различаются по своим аллоферментам не больше, чем две отдельные популяции, принадлежащие к одному виду - D. willistoni. Более того, для того чтобы обнаружить такое же сильное различие по аллоферментам, как между двумя морфологически различными видами группы D. willistoni, нам приходится дойти до вида D. neopicta.
Используя эти значения S, можно построить дендрограмму, аналогичную филогении, построенной по хромосомным данным. Эта филогения, основанная на аллоферментном сходстве, представлена на рис. 3-7 справа. Поразительное сходство между этими двумя дендрограммами подтверждает и усиливает выводы о родственных отношениях, сделанные по результатам изучения инверсий. Так, данные по аллоферментам подтверждают гипотезу относительно обратной миграции D. hemipeza на Оаху с комплекса Молокаи-Мауи. Об этом свидетельствует тот факт, что D. hemipeza находится в более тесном родстве с D. differens и D. plantibia (S равно соответственно 0,74 и 0,78), чем с D. substenoptera (S = 0,40).
На основании этих данных Карсон сумел приложить временной масштаб к скоростям, с которыми происходили изменения ферментов или других белков у этих видов. Это оказалось возможным благодаря наличию точных данных о возрасте островов, на которых обитают эти виды, а также тому обстоятельству, что дрозофилы не могли появиться там до возникновения самих островов. Наиболее подходящая его оценка - накопление 1% генетических различий за 20000 лет. Поскольку эти мухи размножаются довольно медленно (два поколения в год), такая дивергенция происходит примерно за 40000 поколений. Необходимо также обратить внимание на то, что скорость этих изменений, по-видимому, постоянна, а это подтверждает другие наблюдения относительно того, что скорость эволюции белков подчиняется молекулярным часам.
Вариации хромосом и аллоферментов можно использовать для построения схем филогенетических взаимоотношений между этими удивительными существами. А что можно сказать об их морфологии? Эти мухи выделены в самостоятельные виды на основании морфологических различий. Наиболее подходящим примером для наших рассуждений служит сравнение морфологических признаков Drosophila silvestris и D. heteroneura, обитающих на острове Гавайи. Эти два симпатрических вида идентичны по своему кариотипу, имеют одинаковую основную структуру хромосом и сходный тип аллоферментов. Однако они хорошо различаются морфологически. Между ними есть резкие различия в окраске тела и крыльев, а также необычные (для Drosophilidae) различия в форме головы взрослых особей (рис. 3-8).
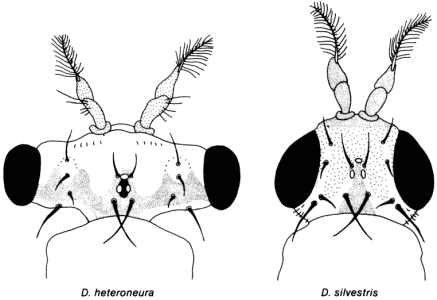
Рис. 3-8. Головы D. heteroneura и D. silvestris со спинной стороны. Эти два вида встречаются симпатрически на большом острове Гавайи - самом молодом из островов Гавайского архипелага. Несмотря на резкие морфологические различия в форме головы между этими двумя видами, они интерфертильны, производя в лабораторных условиях жизнеспособное плодовитое потомство. Как показал генетический анализ этих потомков, различие в форме головы детерминируется всего лишь несколькими генами (Val, 1977).
В природе скрещивание между D. heteroneura и D. silvestris предотвращается эффективным презиготическим изолирующим механизмом, состоящим в специфичном для самок каждого вида сложном ритуале, предшествующем спариванию. Однако в лабораторных условиях действие этого механизма можно преодолеть и получить плодовитые гибриды F1, что позволило Вэл (F. Val) провести реципрокное скрещивание между этими видами и изучить характер наследования морфологических признаков, используемых для их таксономического разделения. По ее оценке, основанной на особенностях наследования таких признаков, как форма и размеры головы, окраска маски, пигментация тела и пятен на крыльях, морфологические различия между D. silvestris и D. heteroneura детерминируются 14-19 локусами. Поскольку геологически остров Гавайи возник не раньше, чем 700000 лет назад, а эти два вида для него эндемичны, их дивергенция не могла произойти раньше этого срока. Различия по аллоферментам, определенные по 25 локусам, ничтожны между данными двумя видами (S = 0,96), так что они дивергировали, по-видимому, даже гораздо позднее, чем возник этот вулканический остров; возможно даже, что их дивергенция произошла всего 70000 лет назад. Если это так, то этот случай видообразования показывает, с какой быстротой может произойти резкое морфологическое изменение.
Для того чтобы показать, что такого рода изменение свойственно не только островной биоте или тропикам, можно привести пример двух симпатрических видов - североамериканских златоглазок Chrysopa carnea и С. downesi, которых изучали Таубер и Таубер (C. Tauber и М. Tauber). Подобно двум рассмотренным выше симпатрическим видам Drosophila, С. carnea и С. downesi в природе изолированы при помощи презиготических механизмов, однако в лаборатории можно добиться их скрещивания, в результате которого они дают плодовитое потомство. Изоляция этих видов в природе обусловлена различиями в их сезонных и суточных ритмах. С. carnea производит три отдельные генерации каждое лето. Осенью взрослые особи третьей генерации впадают в репродуктивную диапазу и перезимовывают в этом состоянии. Что касается С. downesi, то они дают только одну генерацию и размножаются только весной. Остальное время года они находятся в состоянии диапаузы. Характер диапаузы у каждого из этих видов регулируется длиной светового дня. Еще одно различие между ними касается окраски. С. carnea в летние месяцы бывает светло-зеленого цвета, но осенью диапаузирующие насекомые изменяют окраску на красновато-коричневую. С. downesi круглый год остаются темно-зелеными.
Экспериментальные скрещивания, проведенные в лаборатории, показали, что различия между этими двумя видами определяются тремя генами. Два из них регулируют наступление диапаузы в качестве реакции на изменение длины светового дня, а третий - различия в окраске. У нас нет эталона, который позволил бы определить время дивергенции этих двух видов, однако ясно, что степень генетического изменения невелика и что для возникновения такой дивергенции не могло потребоваться много времени.
Ни один из описанных примеров не может служить доказательством того, что видообразование во всех случаях происходит таким образом и за короткие сроки. Они выявляют, однако, два важных момента: видообразование и морфологическая эволюция могут происходить очень быстро и не требуют ни крупных хромосомных перестроек, ни значительных - в количественном аспекте - генетических изменений.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Генетическая грамматика
Генетическая грамматика После установления химического строения и пространственной структуры ДНК оставалось еще множество вопросов, основной из которых заключался в том, как же ДНК кодирует белки, то есть, что представляет из себя генетический код этой молекулы, какую
Глава 8 У порога неоантропов: дивергенция троглодитид и гоминид
Глава 8 У порога неоантропов: дивергенция троглодитид и гоминид I. Характер отбора, лежавшего в основе дивергенцииЕсли ещё раз непредвзято спросить себя об отличительных признаках человека, которые даны опытом истории и не могли бы быть «в другом смысле» распространены
Генетическая грамматика
Генетическая грамматика После установления химического строения и пространственной структуры ДНК оставалось еще множество вопросов, основной из которых заключался в том, как же ДНК кодирует белки, то есть, что представляет из себя генетический код этой молекулы, какую
Знаки препинания (Генетическая пунктуация)
Знаки препинания (Генетическая пунктуация) Как в письменном языке пунктуация (точки, запятые, тире и т.д.) используется для членения и графической организации текста, так и в ДНКовом тексте существуют всевозможные знаки-сигналы, обозначающие границы отдельных
Доступная генетическая вариация
Доступная генетическая вариация Может оказаться, что несмотря на силу возможного давления отбора, эволюция не породит ничего, если нет генетической вариации, над которой она могла бы работать. «Таким образом, хотя можно доказать, что обладание крыльями в дополнение к
Глава 1. Генетическая информация
Глава 1. Генетическая информация Ключевой проблемой биологии, по-видимому, можно считать вопрос о том, как увековечивает свой опыт живая материя. М. Дельбрюк (1906–1981), американский генетик, лауреат Нобелевской премии 1969 г. При изучении любого биологического феномена
Дивергенция признака.
Дивергенция признака. Принцип, который я обозначаю этим термином, крайне важен и, как мне кажется, объясняет некоторые существенные факты. Во-первых, разновидности, даже заметно выраженные и обладающие до некоторой степени сходством с видами, – о чем свидетельствует то
Генетическая детерминация пола
Генетическая детерминация пола Пол — это признак, о котором с наибольшей очевидностью можно сказать, что он наследуется. Несомненно, на развитии признаков пола сказываются и факторы среды: наличие в матке близнеца противоположного пола вызывает у некоторых
11.5. Генетическая психофизиология
11.5. Генетическая психофизиология Генетическая психофизиология изучает взаимодействие генотипа и факторов среды в формировании физиологических систем организма, обеспечивающих психическую деятельность (Булаева К. Б., 1991).Структурная единица нервной системы – нейрон –
Глава 3. Генетическая информация
Глава 3. Генетическая информация Носителями наследственной информации в природе являются нуклеиновые кислоты. Именно они выполняют три необходимые для жизни функции: хранение, воспроизведение и реализацию этой информации. В ходе эволюции ключевая роль по хранению и
Глава 7 Генетическая регуляция развития
Глава 7 Генетическая регуляция развития У природы столько дел в этом мире, ей приходится создавать такую массу разнообразнейших творений, что по временам она и сама не в силах разобраться во всех тех различных процессах, которыми она одновременно занимается. Уилки
Генетическая изменчивость мужских особей
Генетическая изменчивость мужских особей Для чего существуют два пола (рис. 8.39)? Вопрос не так прост, как может показаться на первый взгляд.«Школьное» объяснение заключается в том, что половое размножение дает возможность получать потомство с новыми признаками, которых
Генетическая символика
Генетическая символика Гены обозначаются буквами латинского или греческого алфавита. Доминантные аллели обычно записывают заглавными буквами, а рецессивные – строчными. Иногда символом служат несколько букв – сокращение слова, обозначающее контролируемый признак (vg
3. Генетическая сеть жизни [11]
3. Генетическая сеть жизни [11] Склонись робко перед фактом, словно малое дитя, будь готов оставить любое мнение, любой предрассудок, покорно следуй за природой к любой пропасти — иначе ничего не узнаешь и ничему не научишься. Томас Генри Гексли [12] Когда жарким