Индукция и появление структуры
Индукция и появление структуры
При поверхностном наблюдении за все возрастающей сложностью развивающихся зародышей можно заметить появление хорошо знакомых черт данного животного: различных органов, зубов, конечностей, глаз. Однако эмбриологи нередко оказываются примерно в таком положении, как капитан Ричард, главный герой книги Эрнста Дженджера (Е. Juenger) «Стеклянные пчелы». Столкнувшись с непостижимыми творческими способностями Цаппарони и его хитроумных автоматов, стеклянных пчел, он мог только пробормотать: «Я был вынужден принять эти новые создания такими, какие они есть - я не в состоянии был объяснить их. Почти то же самое мы испытываем, стоя на скале и наблюдая за морскими животными: мы видим рыб и крабов и даже узнаем медуз, но внезапно из глубины всплывают какие-то создания, задающие нам неразрешимые и волнующие загадки».
Одна из таких загадок - очевидный эпигенез, опосредованный индукционными взаимодействиями. Очень большую роль в понимании природы индукции и ее функций в процессе развития сыграли работы Ганса Шпемана (Н. Spemann) и его сотрудников, проведенные после первой мировой войны. Шпеман пересаживал эктодермальные клетки, взятые с поверхности одного зародыша тритона, в поверхностный слой другого такого зародыша. В качестве доноров и реципиентов он использовал виды Triturus, имеющие разную окраску, так что трансплантаты можно было легко отличать от тканей хозяина по наличию или отсутствию пигментных гранул на отдельных участках поверхности химерных зародышей. Если в качестве доноров использовали зародышей на стадии ранней гаструлы, то клетки из разных участков эктодермы развивались в соответствии с дифференцировкой той области реципиента, в которую они были пересажены. Если же донорами служили зародыши, закончившие гаструляцию, то в большинстве случаев наблюдалась иная картина: оказывалось, что пересаженные клетки уже детерминированы, так что, например, презумптивные клетки нейральной эктодермы дифференцировались в нервную ткань независимо от того, в какой участок реципиента они были пересажены. Эти эксперименты дали очень ценные сведения. Они показали, что направление детерминации эктодермальных клеток у амфибий не запрограммировано в самих этих клетках: будет ли данная клетка развиваться в направлении эпителиальной или нервной ткани, зависит от ее местоположения в зародыше во время гаструляции.
Решающий эксперимент - один из самых впечатляющих и значительных для дальнейшего развития эмбриологии - провела ученица Шпемана, Хильде Мангольд (Spemann, Mangold, 1924); она непосредственно показала существование индукционной системы, обусловливающей детерминацию осевых структур. Как это часто бывает с действительно плодотворными работами, экспериментам Мангольд суждено было породить ряд других волнующих загадок.
Как показано на рис. 5-1, во время гаструляции клетки, лежащие на поверхности бластулы амфибии, перемещаются внутрь, проходя через бластопор. В результате образуется двуслойный зародыш. Такая двуслойная организация - необходимая предпосылка для крупных начальных индукционных процессов. Однако, как установили Шпеман и Мангольд, спинная губа бластопора играет некую особую роль. В области спинной губы происходит инвагинация клеток при гаструляции, и, хотя функция этой области до начала гаструляции не проявляется, она обособляется вскоре после оплодотворения, что выражается в образовании серого серпа.
Проведенный Мангольд эксперимент был очень прост: она пересаживала кусочек спинной губы бластопора из ранней гаструлы на боковую поверхность зародыша-реципиента. Однако, в отличие от того, что наблюдалось в более ранних экспериментах с пересадками кусочков эктодермы, пересаженные клетки не развивались в этом случае в соответствии с участком, в который их пересадили. Вместо этого они вызвали образование добавочного набора осевых органов, т.е. получились сиамские близнецы (рис. 5-2). У добавочного эмбриона не было головы (в первых экспериментах Мангольд), но были хорда, парные сомиты и нервная трубка, а также кишка и почечные канальцы. Поскольку для пересадки использовались клетки, отличавшиеся по пигментации от клеток реципиента, можно было установить, какие части добавочного зародыша происходят из пересаженных клеток, а какие - из клеток реципиента. Оказалось, что хорда целиком образована пересаженными клетками, а нервная трубка, сомиты и другие структуры состоят в основном из клеток зародыша-реципиента.
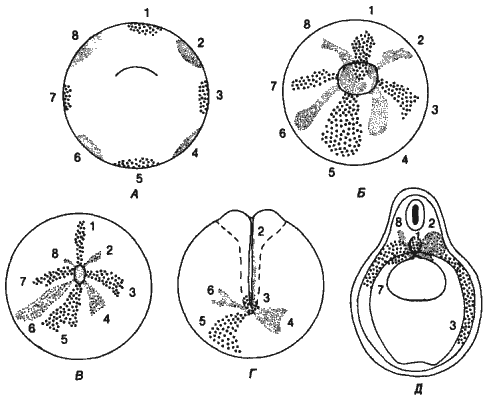
Рис. 5-1. Использование метода маркировки зародыша прижизненными красителями для составления карты перемещений клеток во время гаструляции у жерлянки (Bombinator). А-В. Участки клеток, помеченные прижизненными красителями, втягиваются внутрь зародыша через бластопор. Г. Внешний вид окрашенных зародышей на стадии нейрулы. Д. Поперечный срез зародыша на стадии замыкания нервной трубки; видно распределение окрашенных клеток внутри зародыша. Благодаря этому методу Фогт получил возможность составить первые карты презумптивных зачатков для амфибий (Vogt, 1929).
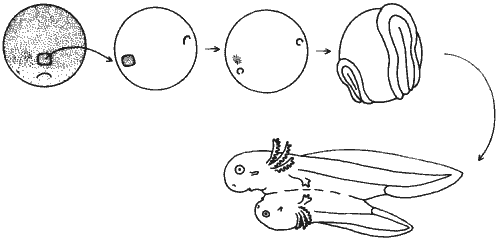
Рис. 5-2. Пересадка клеток спинной губы бластопора от зародыша-донора на брюшную сторону зародыша-реципиента. В результате трансплантации у реципиента образуются добавочные осевые органы (Hadorn, 1974).
Образование большей части тканей, формирующихся в результате пересадки спинной губы, из клеток реципиента показывает, что эти клетки были индуцированы к дифференцировке в измененных направлениях под действием того, чему Шпеман дал название первичного организатора. Это организующее начало находится в клетках спинной губы, которая после инвагинации дает начало крыше первичной кишки, а та в свою очередь, индуцирует образование хорды. Возникновение индуцируемых осевых структур происходит в четко выраженном передне-заднем направлении. Эксперименты с пересадкой кусочков крыши передней кишки показали, что упорядоченность структур центральной нервной системы детерминируется индукторами, которые продуцируются соответствующими участками крыши первичной кишки. Хордомезодерма, действуя снизу вверх, не просто индуцирует в вышележащих тканях развитие нейральных структур: различные участки этой мезодермы продуцируют индукторы, различающиеся по своей информационной специфичности.
В отношении индуцирующих систем можно сделать два обобщения. Индуцирующая ткань должна быть способна вырабатывать необходимое в данном конкретном случае индуцирующее вещество, а ткань-мишень должна обладать соответствующей компетентностью, чтобы реагировать на него. Эти необходимые свойства отражают динамику морфогенетических процессов. Образование индуцирующих веществ и способность реагировать на них - свойства преходящие, и относительное расположение индуцирующей и индуцируемой тканей изменяется по мере течения морфогенеза.
Хрусталик позвоночных - специализированная структура, функция которой состоит в преломлении падающего света. Его развитие начинается с образования презумптивной тканью хрусталика сферического зачатка хрусталика. Затем клетки этого зачатка вытягиваются, превращаясь в волокна хрусталика, которые, ориентируясь определенным образом, формируют преломляющее тело хрусталика. В процессе этих превращений волокна утрачивают ядра и митохондрии, так что хрусталик становится прозрачным. Морфологическая дифференцировка сопровождается клеточной дифференцировкой. Клетки хрусталика, как это пишет в своем обзоре Бломендаль (Bloemendal), начинают синтезировать главным образом группу специализированных белков хрусталика - ?-, ?- и ?-кристаллины. Синтез этих белков, по-видимому, составляет свыше 80% всего белкового синтеза клеток хрусталика и обеспечивается долгоживущими кристаллиновыми мРНК.
Хрусталик образуется из эпидермальных клеток ограниченного участка головы, однако оказалось, что у амфибий можно индуцировать развитие хрусталика из почти любого участка эпидермиса зародыша. Для этого достаточно либо пересадить глазной пузырек в необычное для него место, либо заменить эпидермис, предназначенный для образования хрусталика, кусочком эпидермиса из другой части тела. Глазной пузырь - вырост переднего мозга, дающий начало зрительному нерву и сетчатке, - служит главным, но не единственным, индуктором дифференцировки хрусталика.
Особенно ценные данные о динамичном характере индукционных взаимодействий получил Джекобсон (A. G. Jacobson), исследовавший процесс индукции хрусталика у зародышей амфибий. Морфогенез, происходящий в процессе развития, сопровождается очень значительными перемещениями тканей относительно друг друга. Так, презумптивный эпидермис, из которого развивается хрусталик, во время гаструляции лежит над энтодермальной стенкой будущей глотки, как это показано на рис. 5-3. Эта энтодерма фактически служит первым индуктором хрусталика. По мере дальнейшей гаструляции сердечная мезодерма перемещается вперед, до тех пор пока ее передний край не окажется лежащим под презумптивным хрусталиком и она также не начнет действовать как индуктор. Далее во время нейруляции нервная пластинка замыкается, превращаясь в нервную трубку, и на ее переднем конце начинают выпячиваться глазные пузыри. В результате замыкания нервной трубки клетки будущего хрусталика приходят в соприкосновение с презумптивной сетчаткой, и начиная с этого времени главным индуктором хрусталика служит сетчатка. Степень участия каждой из этих тканей в индукции хрусталика Джекобсон оценивал на основании экспериментов, в которых он удалял ту или иную из индуцирующих тканей, наблюдая при этом за уровнем дифференцировки хрусталика. В экспериментах, в которых сетчатку удаляли, так что индукторами служили только энтодерма и сердечная мезодерма, у 42% зародышей все же формировались хрусталики. Джекобсон пришел к выводу, что в сумме энтодерма и мезодерма равноценны сетчатке в смысле индукции хрусталика. В развитии других органов, таких как нос и ухо (рис. 5-4), также участвует по нескольку индукторов. (Результаты этих и других аналогичных опытов сильно зависят от вида животного и условий эксперимента, и предлагаемая здесь интерпретация не получила пока общего признания.- Прим. ред.)
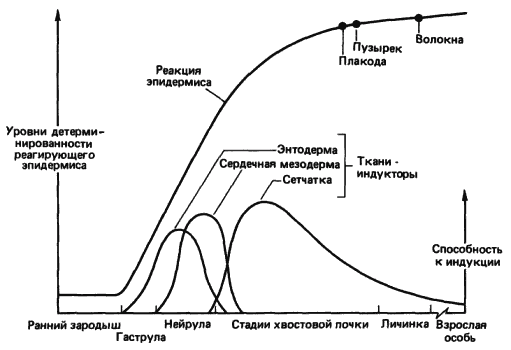
Рис. 5-3. Последовательные индукционные взаимодействия, необходимые для образования хрусталика у зародыша амфибии (Jacobson, 1966).
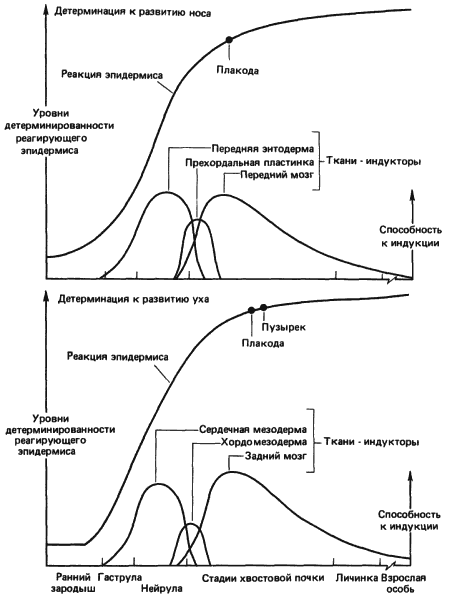
Рис. 5-4. Последовательные индукционные взаимодействия, детерминирующие развитие носа и уха у амфибий (Jacobson, 1966).
Сети индукторов могут играть важную роль в канализации развития, обеспечивая нормальное течение органогенеза, даже если один из компонентов индуцирующей системы не сумеет произвести сигнал нужной силы. И, как понял Джекобсон, многочисленность индуцирующих тканей может иметь решающее значение для точного установления места формирования данного органа. Джекобсон провел ряд экспериментов, в которых либо полоску головной эктодермы, образующую в конечном итоге нос, хрусталик и ухо, либо лежащую под ней нервную пластинку поворачивали на 180° на разных стадиях развития. Результаты этих экспериментов ясно показали, что, хотя головной мозг оказывает наиболее сильное индуцирующее действие, необходимое для полного развития этих структур, местоположение носа, хрусталика и уха действительно частично детерминируется более ранними индукционными событиями при участии энтодермы и проспективной сердечной мезодермы.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
ПОЯВЛЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ
ПОЯВЛЕНИЕ ЦВЕТКОВЫХ РАСТЕНИЙ Шишка, или цветок беннеттита (слева), ицветок магнолии (часть органов растенийудалена)В мезозойскую эру произошли два великих события в истории жизни на Земле, которые изменили и обновили органический мир Земли: появились первые
ПОЯВЛЕНИЕ ЧЕЛОВЕКА
ПОЯВЛЕНИЕ ЧЕЛОВЕКА Человек — это звучит гордо! М.
3. Индукция
3. Индукция В «классической» механике развития эмбриональной индукцией называют такое влияние одной ткани на другую, соседнюю, которое вызывает в месте контакта новую дифференцировку. Иногда (обычно в искусственной экспериментальной ситуации) индуцирующая ткань
8.1. Появление эукариот
8.1. Появление эукариот Ранее мы обсуждали вопросы развития биотического круговорота на основе прокариот. Оказалось, что главные достижения в совершенствовании обеих ветвей цикла как синтетической, так и деструктивной связаны с улучшением энергетики, повышением
§ 32. Появление конечностей
§ 32. Появление конечностей На первый взгляд происхождение конечностей наземных позвоночных не вызывает каких-либо логических проблем. Их появление вторично по отношению к миграции предков наземных позвоночных на мелководье и формированию эффективного воздушного
ОТ СТРУКТУРЫ К ФУНКЦИИ (функциональная геномика)
ОТ СТРУКТУРЫ К ФУНКЦИИ (функциональная геномика) Недостаточно знания, необходимо также применение; недостаточно хотеть, надо и делать. И. Гёте Хорошо известно выражение, что от слова «халва» во рту слаще не становится. Так же обстоит дело и с нашим геномом. В нем есть много
Появление человека
Появление человека В Европе и Азии, в том числе и в нашей стране, учёные обнаружили громадное скопление костей — целые «кладбища» животных, живших несколько миллионов лет назад. Они раскопали многочисленные кости антилоп, газелей, жирафов, гиен, тигров, обезьян и других
1. ПОЯВЛЕНИЕ ПСИХИЧЕСКОГО
1. ПОЯВЛЕНИЕ ПСИХИЧЕСКОГО Наиболее принятой в настоящее время точкой зрения является та, что психическое – атрибут живых систем, а неживая природа, включая созданные людьми сложные устройства, психикой не обладает. Это важное отличие живых систем от неживых отмечено П.
Полуавтономные структуры
Полуавтономные структуры Помимо ядра, небольшая доля генетической информации клетки находится в таких органоидах, как митохондрии и хлоропласты, имеющих собственные генетические системы. Как митохондрии, так и хлоропласты имеют свои ДНК, все виды РНК (и-РНК, т-РНК, р-РНК)
I. ВАРИАЦИИ ОКРАСОВ И СТРУКТУРЫ ШЕРСТИ
I. ВАРИАЦИИ ОКРАСОВ И СТРУКТУРЫ ШЕРСТИ Прежде, чем приступить к изложению генетики пород собак, необходимо в общих чертах ознакомиться с тем, что известно о генах, определяющих окрасы и структуру шерсти у собак, и их действии. Для этого было бы полезно иметь представление
4.3. Морфогенетические поля как вероятностные структуры
4.3. Морфогенетические поля как вероятностные структуры Орбитали электронов вокруг атомного ядра можно рассматривать как структуры в морфогенетическом поле атома. Эти орбитали могут быть описаны решениями уравнения Шредингера. Однако, согласно квантовой механике,
Появление креодонтов
Появление креодонтов Когда приоткрываются двери прошлого, мы видим устремленные на нас бессмысленные взоры чудовищ. У этих фантастических порождений прошлой жизни размеры тела намного опережали развитие их нервной организации.На страницах нашей книги уже находили
Изучение структуры популяции
Изучение структуры популяции Выше неоднократно подчеркивалась важность точной характеристики элементарной эволюционной единицы — популяции. Одна из самых важных популяционных характеристик — пространственная структура популяции. От того, на какие группы особей