Сколько генов необходимо для развития?
Сколько генов необходимо для развития?
К счастью, существуют методы, позволяющие оценить количество генетической информации, имеющейся у высших организмов. Один из самых тонких таких методов - классическая менделевская генетика. Главная трудность, связанная с этим методом, заключается в удивительно малом числе организмов, которые достаточно подробно изучены генетически, чтобы предоставить нам желаемые данные. В сущности, на сегодняшний день лишь один организм - плодовую мушку Drosophila melanogaster - можно считать достаточно хорошо изученной генетически. С тех пор как в 1910 г. Т. Морган и его ученики впервые внесли дрозофилу в лабораторию, по одному этому скромному насекомому было собрано внушительное количество информации. С помощью химических мутагенов и рентгеновых лучей у дрозофилы были индуцированы многие тысячи мутаций, разбросанных по всему ее геному. Современный каталог мутаций D. melanogaster - свидетельство усердия генетиков - занимает свыше 500 страниц.
Эта обширная коллекция мутаций, а также умение получать новые создают возможность для выявления зависимости между генами и морфологическими, метаболическими, морфогенетическими или поведенческими признаками организма. Мутации можно использовать для получения двух основных типов информации относительно развития: это, во-первых, важнейшие сведения относительно функции отдельных процессов онтогенеза и их регуляции, получаемые путем анализа способов, которыми мутации вмешиваются в такие процессы; во-вторых, это информация (она нас, собственно, здесь и интересует) о числе генов, контролирующих процессы развития, которая в свою очередь дает нам возможность оценить долю генома, участвующего в онтогенезе как таковом.
Каждый отдельный ген может дать ряд различных мутантных аллелей, проявляющихся в виде различных фенотипов; поэтому число генов, участвующих в том или ином процессе, нельзя оценить с помощью такой привлекательной по своей простоте процедуры, как подсчет мутаций. Рассмотрим две мутации, воздействующие на один определенный фенотипический признак. Вопрос о том, относятся ли эти две мутации к одному или к двум генам, решается при помощи стандартного теста на комплементацию.
Число групп комплементации, регулирующих развитие дрозофилы, можно оценить, взяв выборку мутаций какого-либо одного класса и определяя путем экстраполяции общее число генов этого класса во всем геноме. Такая экстраполяция основана на двух допущениях. Первое из них состоит в том, что гены, имеющие одинаковые функции, распределены либо в одной хромосоме, либо во всем геноме случайным образом; т.е. гены, выполняющие в развитии близкие функции, не образуют кластеров. За некоторыми важными исключениями, рассмотренными в гл. 8, это допущение кажется разумным. Второе допущение заключается в том, что гены, несущие сходные функции, мутируют с одинаковой частотой и что их мутации одинаково легко выявить. Это допущение не выполняется отдельными генами, которые оказываются более мутабильными, чем большинство других, но оно, по-видимому, оправданно применительно к большой выборке генов. Можно выявить мутации, блокирующие развитие различными способами, и сгруппировать их в определенные категории. По каждой категории собирают как можно больше мутаций, но сделать исчерпывающую выборку, в которой геном насыщен мутантными генами желаемой категории, конечно, невозможно. Поэтому берется ненасыщающая выборка, которую надо затем статистически расширить. Для этого необходимо, чтобы гены, попавшие в выборку просто потому, что они мутировали, действительно представляли собой случайную выборку из того класса, который интересует исследователя.
Какие классы генов исследователь считает существенными для развития, зависит от особенностей развития изучаемого организма. Важно помнить, что дрозофила - голометаболическое насекомое, т.е. ее развитие сопровождается полным превращением, и личинка по своей морфологии, физиологии и поведению совершенно не похожа на взрослую муху. При этом личиночные структуры не превращаются непосредственно в соответствующие имагинальные структуры. Муха - это своего рода феникс: большая часть ее личиночных тканей распадается и поглощается тканями взрослой особи, развивающимися из имагинальных дисков, находящихся в личинке. Таким образом, в этом организме на самом деле имеются две морфогенетические системы, в которых может осуществляться генная функция, играющая решающую роль в развитии взрослой особи. Первая система действует во время эмбриогенеза, когда происходит образование составных частей и морфологии личинки. Вторая система определяет становление, поддержание и пролиферацию имагинальных дисков в личинке и их дифференцировку, приводящую к формированию взрослой особи.
Первая из этих систем - личиночное развитие - состоит в действительности из двух разделенных во времени систем действия генов. Содержащаяся в яйце информация, необходимая для ранних стадий развития, представляет собой продукты действия генов во время оогенеза. По прошествии этого периода зависимости от генных продуктов, созданных во время оогенеза, в зародыше начинается активная транскрипция, поставляющая информацию, необходимую для дальнейшего развития. Таким образом, у дрозофилы в набор важных для развития генов должны входить гены, наследуемые по материнскому типу (т.е. функционирующие во время оогенеза), гены, функция которых имеет жизненно важное значение в течение зародышевого или личиночного развития, и гены, специфически воздействующие на развитие имагинальных дисков. Для дрозофилы сделаны оценки генов всех этих трех классов.
Ганз (Gans) и его сотрудники, а также Молер (Moller) выделили большое число мутаций с материнским эффектом для генов, локализованных в Х-хромосоме (обсуждение хромосом дрозофилы см. гл. 3), а Раис и Герен (Rice, Garen) выделили такие мутации, локализованные в третьей хромосоме. Существуют два типа мутаций с материнским эффектом. В первом случае самки производят морфологически аномальные яйца (например, с дефектами яйцевой оболочки), а во втором - яйца, которые кажутся нормальными, но не могут завершить развитие. Нас интересуют только мутации этого последнего класса. Как видно из табл. 10-1, Ганз выявил 42 мутации этого класса, Молер - 146, а Раис и Герен - 6. Проведенные затем тесты на генетическую комплементацию показали, что все эти мутации распадаются на 30, 60 и 5 отдельных групп комплементации соответственно, т. е. некоторые из этих мутаций возникли в результате повторных попаданий в один и тот же ген.
Таблица 10-1. Оценки общего числа генов с материнским эффектом в геноме дрозофилы
Авторы Хромосома Общее число мутаций Число мутировавших генов n0 Оценка общего числа генов Ганз X 42 30 53 83 Молер X 146 60 38 98 Раис и Герен Третья 6 5 8 13 Результаты экстраполяции Все — — — 117Прежде чем приступить к анализу этих мутаций, следует напомнить предостережение, высказанное Томасом Гексли: «Математику можно сравнить с наисовершеннейшей мельницей, которая размелет ваш продукт в самую тонкую муку, но при этом то, что вы получите, зависит от того, что вы положите на жернова...». Используя допущения о случайном размещении генов и их одинаковой мутабильности, число немутировавших генов можно оценить при помощи распределения Пуассона. При этом исходят из допущения высокой вероятности того, что у большинства генов данного класса не возникло ни одной мутации, у небольшого числа возникло по одной мутации, а вероятность двух мутаций на ген еще меньше. Количественная зависимость между группами генов с отсутствием мутаций, одной и двумя мутациями описывается выражением
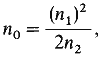
где n1 - число генов, мутировавших один раз, n2 - число генов, мутировавших дважды, и n0 - число генов, у которых не обнаружено ни одной мутации. Численные значения для n1 и n2 можно получить по данным комплементации для любого класса.
На основании данных Ганза и его сотрудников n0 получается равным 53, а по данным Молера п0 равно 38 при оценках общего числа генов в Х-хромосоме, наследуемых по материнскому типу, равных 83 и 98 соответственно. С учетом характера оценок совпадение двух последних чисел следует считать очень хорошим; можно принять, что в Х-хромосоме имеется примерно 90 генов, жизненно важных для эмбриогенеза, и что продукты этих генов образуются в результате их активности во время оогенеза. Точно так же из данных Раиса и Герена можно получить n0=8 и nобщее=13 для генов этого класса, локализованных в третьей хромосоме. Расхождения между оценками для Х-хромосомы и третьей хромосомы интересны, но объяснить их нелегко, потому что третья хромосома содержит примерно вдвое больше ДНК, чем Х-хромосома, и такое несовпадение нельзя отнести за счет различий в величине изученных выборок. Поэтому создается впечатление, что гены с материнским эффектом находятся преимущественно в Х-хромосоме.
Если допустить, что вторая хромосома - аутосома, сходная по содержанию ДНК с третьей хромосомой, - сходна с ней также и по своей организации, то в ней также, вероятно, имеется 13 генов с материнским эффектом. Последняя аутосома - очень маленькая четвертая хромосома - судя по ее относительным размерам, должна содержать один ген с материнским эффектом. Если суммировать все эти оценки, то получится, что в геноме дрозофилы имеется в общей сложности примерно 117 генов с материнским эффектом, причем 90 из них локализованы в Х-хромосоме и, судя по результатам изучения третьей хромосомы, - еще примерно 27 таких генов локализовано в аутосомах.
Для того чтобы оценить число генов, необходимых для зародышевого развития, но не относящихся к классу генов с материнским эффектом, нам придется прибегнуть к несколько иному методу. Можно взять случайную выборку летальных мутаций и определить число тех из них, которые детальны в период зародышевого развития. Затем путем экстраполяции можно установить общее число таких деталей во всем геноме, используя долю эмбриональных деталей среди всех летальных мутаций в случайной выборке для определения общего числа генов в данном геноме, способных давать летальные мутации. Мы здесь и пользуемся этим методом, с той разницей что наши случайные выборки летальных мутаций были взяты из двух маленьких, но репрезентативных участков генома, насыщенных летальными мутациями.
Первое требование состоит, конечно, в том, чтобы получить оценку общего числа генов в геноме дрозофилы. Джадд (Judd) и его сотрудники в результате интенсивного генетического анализа небольшого участка Х-хромосомы установили, что число полос в политенных хромосомах дрозофилы почти соответствует числу генов, определенному методом генетической комплементации. Общее число полос, различимых на политенных хромосомах Drosophila melanogaster (рис. 10-1), равно 5000, а по разумной оценке общее число генов также равно 5000. Излишне говорить, что с тех пор как в 1972 г. была впервые предложена эта оценка, она горячо оспаривалась генетиками, но дальнейшие исследования продолжали подтверждать примерное соответствие между числом полос и числом генов.
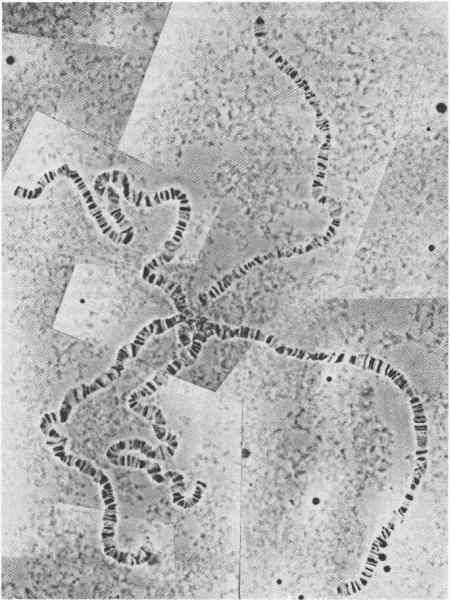
Рис. 10-1. Микрофотография политенных хромосом слюнной железы Drosophila melanogaster (фотография любезно предоставлена R. Lewis).
Доля этих 5000 генов, способных давать летальные мутации, была определена в двух работах: Шеннон (Shannon) и ее сотрудников и Хочмена (Hochman). Шеннон и др. исследовали мутации в небольшом участке Х-хромосомы между генами zeste и white, который, как показано на рис. 10-2, содержит 13 полос и 13 генов, поддающихся идентификации. Хочмен изучил подобным же образом маленькую четвертую хромосому, содержащую всего 50 полос, в которой он идентифицировал 43 гена. Из 13 генов, локализованных в участке Х-хромосомы, изученном Шенноном и др., только 2 не дают летальных мутаций, а из 43 генов четвертой хромосомы, выявленных Хочменом, - только 6, т.е., по-видимому, эти гены не имеют жизненно важного значения для дрозофилы. На основании экстраполяции от этой ограниченной выборки можно считать, что примерно 85% из 5000 генов дрозофилы способны давать летальные мутации.
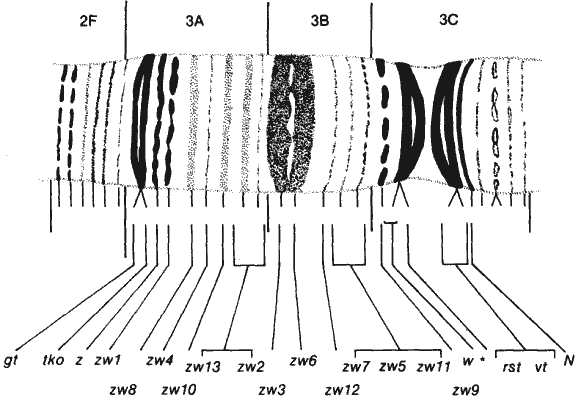
Рис. 10-2. Идентификация генов, локализованных в небольшом участке X-хромосомы Drosophila melanogaster, расположенном между генами zeste и white. Тринадцать идентифицированных генов соответствуют тринадцати видимым дискам (Kauffman et al., 1975; с изменениями).
Подвергнув эти обнаруженные и нанесенные на карты летали морфогенетическому анализу, Шеннон и др. и Хочмен установили, что 1/10 и 5/37 соответственно были эмбриональными деталями. Если относительная частота эмбриональных деталей среди всех деталей точно отражает общее их число в геноме, то в таком случае, как это суммировано в табл. 10-2, у дрозофилы имеется от 425 до 550, или примерно 500 генов, способных давать мутации не просто летальные, но летальные именно во время зародышевого развития. Суммируя эту оценку с оценкой числа деталей с материнским эффектом, получаем, что для ранних стадий эмбриогенеза и для формирования функционирующей личинки необходимо 617 генов.
Таблица 10-2. Оценка общего числа эмбриональных леталей в геноме дрозофилы
Авторы Участок Число полос Число генов Число леталей Число эмбриональных леталей Число эмбриональных леталей во всем геноме Шеннон и др. zeste white 13 13 10 1 425 Хочмен Четвертая хромосома 50 43 37 5 550Оценку числа генов, необходимых для развития и поддержания имагинальных дисков при отсутствии у зародыша каких-либо дефектов, можно произвести на основании результатов, полученных Ширном и Гереном (Shearn, Geren). Эти исследователи выбрали летальные мутации, которые допускали внешне нормальный эмбриогенез и личиночное развитие, но вызывали гибель во время метаморфоза. Было установлено, что эти мутации обусловливают либо дефекты имагинальных дисков, либо полное их отсутствие. Всего было выделено из третьей хромосомы 57 мутаций такого типа. Тесты на комплементацию среди этих 57 мутаций выявили 52 группы комплементации, или гена. Тем же способом, который использовался для подсчета генов с материнским эффектом, было установлено, что для генов этого класса в третьей хромосоме n0 = 384. Таким образом, теоретически общее число генов в третьей хромосоме, функция которых необходима для нормального развития имагинальных дисков, равна 436. Кисе (Kiss) и его сотрудники обнаружили мутации этого класса также в Х-хромосоме. Поскольку эти мутации представляют собой сцепленные с полом летали, которые нельзя подвергнуть стандартному тесту на комплементацию, Кисе и сотр. были вынуждены провести более сложный анализ, описывать который здесь нет необходимости. По полученной ими оценке для развития функции имагинальных дисков жизненно необходимо 118 генов, локализованных в Х-хромосоме. Если допустить, что во второй и четвертой хромосомах, судя по содержанию в них ДНК по сравнению с третьей хромосомой, локализовано 436 и 20 таких генов соответственно, то общее число генов во всем геноме, необходимое для развития имагинальных дисков, составит приблизительно 1000.
Итак, в целом для нормального течения эмбриогенеза и метаморфоза необходимо 1617 генов, т. е. примерно 30% общего числа генов, определяемого методами менделевской генетики.
Следует подчеркнуть, что в эти оценки совершенно не входят гены некоторых типов - гены, существующие в множественных копиях, такие как гены гистонов, рибосомных РНК (рРНК) и транспортных РНК (тРНК), которые обсуждаются далее в этой главе. Рассмотренными нами способами обнаружения мутаций вряд ли можно выявить гены, существующие в нескольких идентичных копиях. Кроме того, наши оценки, по-видимому, не учитывают гены, определяющие жизненно важные (housekeeping) функции мухи на протяжении всей ее жизни. Так, например, действие мутации, затрагивающей синтез цитохрома с, не будет обладать временной специфичностью, подобно рассмотренным выше мутациям. Если все эти рассуждения верны, то в соответствии с оценкой, полученной путем отбора генов, необходимых для развития, вероятно, можно считать, что у дрозофилы генетическая стоимость развития от яйца до взрослой мухи составляет 30% общего числа генов в геноме.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
1. Промоторы генов
1. Промоторы генов В этом разделе мы кратко расскажем о том, какие нуклеотидные последовательности, прилегающие к генам, а иногда и внутри гена, ответственны за процесс транскрипции. У прокариот эти участки, с которыми связывается молекула РНК-полимеразы и откуда
Сколько же всего генов у человека?
Сколько же всего генов у человека? Это наиболее интересный вопрос, ради которого собственно и затевалось полное секвенирование генома человека. После получения основной информации о структуре генома человека в первую очередь были произведены различные анализы по
Сколько же всего генов у человека?
Сколько же всего генов у человека? Это наиболее интересный вопрос, ради которого собственно и затевалось полное секвенирование генома человека. После получения основной информации о структуре генома человека в первую очередь были произведены различные анализы по
Дупликация генов
Дупликация генов МНОГОФУНКЦИОНАЛЬНЫЕ ГЕНЫ — ОСНОВА ЭВОЛЮЦИОННЫХ НОВШЕСТВ.Мысль о том, что дупликация генов служит важнейшим источником эволюционных новшеств, была высказана еще в 1930-е годы выдающимся биологом Джоном Холдейном (Haldane, 1933). Сегодня в этом нет никаких
В поисках генов видообразования
В поисках генов видообразования Итак, постзиготическая изоляция возникает в результате фиксации несовместимых (конфликтующих) аллелей в разобщенных популяциях. Что же это за гены, изменения которых порождают такие конфликты? В принципе при скрещивании представителей
4.3. Взаимодействие генов
4.3. Взаимодействие генов В организме одновременно функционирует множество генов. В процессах реализации генетической информации в признак возможны многочисленные «пункты» взаимодействия разных генов на уровне биохимических реакций. Такие взаимодействия неизбежно
7.1. Выделение генов
7.1. Выделение генов Возможно использование нескольких путей выделения генов. Каждый из них имеет свои достоинства и недостатки.Химический синтез генов, т. е. синтез нуклеотидов с заданной последовательностью, соответствующей одному гену, впервые был осуществлен в
ДРЕЙФ ГЕНОВ
ДРЕЙФ ГЕНОВ Иногда эта концепция называется «эффект Сьюэлла — Райта», в честь предложивших ее двух популяционных генетиков. После того как Мендель доказал, что гены являются единицами наследственности, а Харди и Вайнберг продемонстрировали механизм их поведения,
Дифференциальная экспрессия генов в процессе развития
Дифференциальная экспрессия генов в процессе развития Один из главных и общепризнанных догматов современной эмбриологии состоит в том, что, за исключением нескольких особых случаев, все клетки данного организма, независимо от того какими они становятся в
Глава 10 Адаптации экспрессии генов в процессе развития
Глава 10 Адаптации экспрессии генов в процессе развития Жизнь -это сила, которая проделывает бесчисленное множество экспериментов, пытаясь организовать себя ... мамонт и человек, мышь и мегатерий, мухи и отцы церкви - все это результаты более или менее успешных попыток
О чем необходимо помнить владельцу собаки, больной эпилепсией
О чем необходимо помнить владельцу собаки, больной эпилепсией Как уже указывалось, лечение больной собаки проводит ветеринарный врач, только он может назначать препарат в определенных дозах. Уменьшать дозы надо постепенно, нельзя делать перерыв в курсе лечения. Если
Глава 15. Это необходимо знать
Глава 15. Это необходимо знать Как дать собаке лекарство Способы введения лекарственных препаратов можно разделить на добровольные и насильственные.Добровольные методы применяются при сохранении у больного животного аппетита. Лекарство дается собакам с пищей. Таким
5. Теория стадийного развития и особенности развития животных
5. Теория стадийного развития и особенности развития животных В основе управления развитием организмов лежит теория стадийного развития, которую сформулировал академик Т. Д. Лысенко, исходя из работы И. В. Мичурина и многочисленных собственных исследований.Несмотря на
Сколько генов у человека
Сколько генов у человека Каждый год элита молекулярной биологии собирается на симпозиум в городке Колд-Спринг-Харбор на живописном северном побережье Лонг-Айленда, что неподалеку от Нью-Йорка. В мае 2000 года там обсуждалось — иначе и быть не могло! — секвенирование