Изменения в организации яйца при возникновении эволюционно продвинутых групп первичноротых
Изменения в организации яйца при возникновении эволюционно продвинутых групп первичноротых
Среди Spiralia наблюдается несколько довольно любопытных крупных эволюционных направлений, затрагивающих ранние стадии развития. Одно из них - это сочетание невероятно консервативного спирального дробления с чрезвычайно разнообразными планами строения тела взрослых особей, обнаруженное у представителей нескольких типов. Так, если обратиться к рассмотренным нами группам, у кольчецов (с их высокометамерным строением тела) мало общего с несегментированными моллюсками. Однако и те и другие обладают не только одинаковым типом дробления, но и сходной судьбой отдельных клеток. Так, из первого квартета микромеров образуется личиночная эктодерма; некоторые микромеры второго квартета образуют эктомезодерму; макромеры - энтодерму, а клетка 4d в конечном счете дает дефинитивную мезодерму взрослой особи. Во многих случаях консервативный способ дробления может быть связан с тем, что у большинства Spiralia личинки ведут планктонный образ жизни: одним из основных признаков всех типов, относящихся к Spiralia, является трохофора или сходная с ней личинка. Эволюционные модификации дробления очень легко коррелировать с утратой или изменением какой-либо структуры на личиночных стадиях, как это было описано выше на примере кольчеца Tubifex или моллюска Unio.
Другим крупным эволюционным направлением среди Spiralia было наблюдаемое в некоторых группах радикальное отклонение ранних стадий дробления от традиционного спирального типа. Это произошло у моллюсков при возникновении головоногих, которые, несмотря на высокое развитие нервной системы, органов чувств и локомоторной системы, сохраняют план строения тела, типичный для моллюсков. Головоногие продуцируют очень крупные яйца, богатые желтком, из которых путем прямого развития без выраженной личиночной стадии, образуются взрослые особи. Дробление ограничено тонким слоем цитоплазмы на поверхности яйца и, как подчеркивает Арнольд (Arnold), совершенно не похоже на дробление у Spiralia. Изменения в типе дробления и морфогенезе, наблюдаемые у головоногих, сходны с изменениями, произошедшими у костистых рыб и птиц, у которых ранние процессы развития также адаптированы к наличию в яйце очень большого количества желтка.
Аналогией возможному ходу изменений в эволюции яйца у головоногих служат высшие кольчецы с их крупными, богатыми желтком яйцами и прямым развитием. Дробление у этих форм модифицировано и большую часть массы яйца составляют альбуминотрофные клетки, несущие исключительно питательную функцию.
Подобно головоногим, членистоногие происходят от предков, принадлежавших к Spiralia, но характер их развития сильно модифицировался. Членистоногих обычно объединяют в один тип, близкородственный кольчецам. Как для кольчецов, так и для членистоногих характерны строгая метамерия в строении тела, ряды повторяющихся придатков, брюшная нервная цепочка и расположенное дорсально сердце. Однако у трех главных групп ныне живущих членистоногих - Crustacea (креветки, морские желуди и т.п.), Chelicerata (мечехвосты, пауки и т.п.) и Uniramia (онихофоры, многоножки и насекомые) - имеется ряд признаков, на основании которых современные исследователи филогении членистоногих (Manton, D. Anderson и Cisne) рассматривают их как искусственную группу. В соответствии с их представлениями Crustacea, Chelicerata и Uniramia возведены в ранг отдельных типов, возникших независимо один от другого.
Ракообразные-единственные членистоногие, у которых сохранилось спиральное дробление. На рис. 4-14 показано дробление у усоногого рачка Tetraclita. Бластомеры перенумерованы в соответствии с системой обозначений, которую предложил Андерсон (Anderson) для бластомеров у Spiralia, и с его представлениями о возможных гомологиях между этими клетками и клетками зародышей Spiralia. Ценность подобного сопоставления типов дробления, по-видимому, ограничена, потому что дробление у ракообразных так глубоко отличается от классического спирального дробления, что любая попытка провести какую-либо аналогию будет безуспешной.
Однако карты с указанием судьбы отдельных участков поверхности бластулы ракообразных можно сравнить с аналогичными картами для кольчецов, как это сделал Андерсон. Как показывает рис. 4-15, на котором представлены две такие карты, у кольчецов презумптивная мезодерма лежит позади презумптивной средней кишки, тогда как у зародышей ракообразных она лежит между средней кишкой и презумптивным первичным ртом. Это изменение касается не только типа дробления, но и основных взаимоотношений между различными участками яйца.
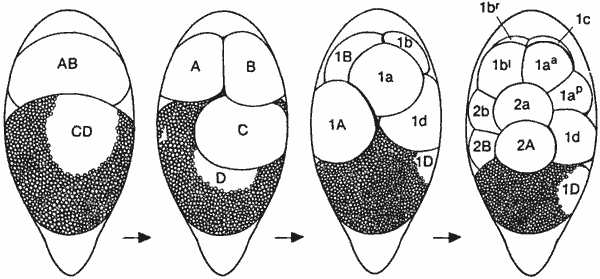
Рис. 4-14. Дробление у усоногого рачка Tetraclita как пример сохранения у ракообразных спирального дробления в сильно модифицированном виде (Anderson, 1969).
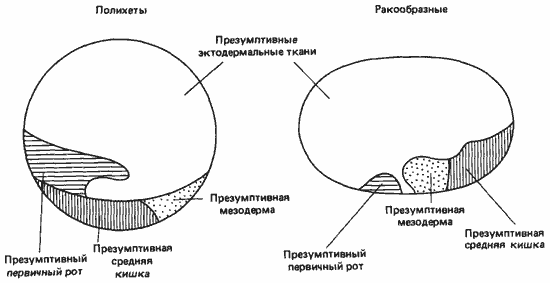
Рис. 4-15. Карта зачатков у зародышей полихет и ракообразных на стадии бластулы (Anderson, 1973).
Большинство других членистоногих, не относящихся к ракообразным, производят яйца, очень богатые желтком; в этих яйцах желточная масса не делится, оставаясь в виде неразделившегося синцития, содержащего образующиеся при дроблении ядра. По окончании делений дробления эти ядра мигрируют к поверхности яйца, где в результате целлюляризации они образуют бластодерму. Прямое сравнение подобного способа деления со спиральным дроблением, разумеется, невозможно. Тем не менее Андерсон сравнивал карты зачатков таких членистоногих с картами зачатков кольчецов. Онихофоры - самая примитивная группа среди Uniramia; в сущности, они так примитивны, что их часто выделяют в отдельный тип, промежуточный между кольчецами и членистоногими. Эти животные обладают некоторыми признаками кольчецов, в частности мягкими покровами и в основном однородными недифференцированными сегментами, а мышечные слои расположены у них так же, как у кольчецов. При этом, однако, у онихофор имеются некоторые признаки членистоногих, например челюсти, ходильные ноги, снабженные коготками, и кровеносная система, сходная с аналогичной системой членистоногих. Наличие трахеи указывает на их близость к насекомым и многоножкам. Сравнение карты зачатков онихофор и кольчецов привело к интересному результату: оказалось, что в отличие от ракообразных карта зачатков зародыша онихофор сходна с картой зародыша кольчецов. Поэтому представляется вероятным, что ракообразные и Uniramia возникли независимо друг от друга. Происхождение этих двух групп было связано с модификацией яйца предкового Spiralia в двух совершенно различных направлениях. У ракообразных сохранилось спиральное дробление, но оно полностью модифицировалось и судьба различных участков зародыша изменилась. Uniramia совершенно отказались от спирального дробления и перешли к синцитиальному типу дробления, предшествующему образованию бластодермы, как адаптации к очень крупным, богатым желтком яйцам. Но судьба отдельных участков яйца сохранила сходство с их судьбой у кольчецов.
Считается, что насекомые, наиболее продвинувшиеся в эволюционном отношении среди всех первичноротых, произошли от какого-то предка, сходного с кольчецами, пройдя через стадию сходства с онихофорами, а затем с многоножками. В процессе такой эволюции постепенно возникали характерное для Uniramia строение ноги и головы, а также специализация сегментов и уменьшение их числа. Морфогенетические и генетические события, лежащие в основе этих изменений, рассматриваются в гл. 7 - 9.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
7. Созревание яйца
7. Созревание яйца Ооцит, достигший больших размеров, еще не яйцо. Для того чтобы стать яйцеклеткой, способной к оплодотворению и дальнейшему развитию, он должен пройти созревание. Это кратковременный, но важный процесс, так как в ходе созревания ооцит приобретает целый
1. Что происходит при возникновении различий
1. Что происходит при возникновении различий В 20-е годы немецкий ученый Фогт сумел проследить судьбу разных частей яйца в развитии. Для этого он окрашивал небольшие участки бластулы амфибий безвредными витальными красителями и наблюдал движение этих участков во время
ПЕРВЫЕ МЛЕКОПИТАЮЩИЕ СОВРЕМЕННЫХ НАМ ГРУПП
ПЕРВЫЕ МЛЕКОПИТАЮЩИЕ СОВРЕМЕННЫХ НАМ ГРУПП Мы говорили, что к концу мезозоя на Земле уже сформировались почти все группы млекопитающих, которые живут и сегодня. Прогрессивное развитие мозга, теплокровность и живорождение сделали их к этому времени более
4.2. Возрастные изменения морфофункциональной организации нейрона
4.2. Возрастные изменения морфофункциональной организации нейрона На ранних стадиях эмбрионального развития нервная клетка имеет большое ядро, окруженное незначительным количеством цитоплазмы. В процессе развития относительный объем ядра уменьшается. Рост аксона
О внезапном появлении целых групп родственных видов.
О внезапном появлении целых групп родственных видов. Многие палеонтологи, например Агассиц, Пикте и Седжвик (Sedgwick), настойчиво указывали на внезапное появление в некоторых формациях целых групп видов как на фатальное возражение против идеи о трансмутации видов. Если бы
3 От яйца до взрослого
3 От яйца до взрослого О, для чего господь, создатель мудрый, Велевший, чтобы жили в небесах Одни лишь духи мужеского пола, Такую новость на Земле устроил, Природы обольстительный порок, Не создал здесь одних мужчин без женщин, И способа иного не нашел Производить
Глава III ОТ ГРУПП КРОВИ К ГЕНАМ
Глава III ОТ ГРУПП КРОВИ К ГЕНАМ Трудно найти в облике человека черту более характерную, чем волосы. О них в первую очередь упоминают, описывая внешность появившегося на свет младенца, нового знакомого или давая приметы преступника. Темные или светлые, волнистые или прямые,
Специфика отбора, организации и методика проведения детских групп
Специфика отбора, организации и методика проведения детских групп Итак, в чем же секрет эффективности метода? Он прост: дети взаимодействуют с животными в эмоционально позитивной и комфортной обстановке. Они просто играют, играют в хендлеров (специалист-кинолог, который
Специфика отбора, организации и методика проведения детских групп
Специфика отбора, организации и методика проведения детских групп Итак, в чем же секрет эффективности метода? Он прост: дети взаимодействуют с животными в эмоционально позитивной и комфортной обстановке. Они просто играют, играют в хендлеров (специалист-кинолог, который
Эволюционные изменения в организации яиц со спиральным дроблением
Эволюционные изменения в организации яиц со спиральным дроблением Для многих зародышей со спиральным дроблением, в особенности для зародышей моллюсков и кольчецов, составлены очень точные карты, на которых указана дальнейшая судьба различных клеток. Разные типы
Эволюционные изменения в организации яиц хордовых
Эволюционные изменения в организации яиц хордовых Для Spiralia характерно развитие, при котором отдельные бластомеры на ранних стадиях дробления уже запрограммированы к дифференцировке в определенном направлении, не изменяющемся даже в том случае, если изолировать их от
8.4.5. Прицельная трансформация групп клеток мозга
8.4.5. Прицельная трансформация групп клеток мозга Были проведены также эксперименты по ненаследуемому повышению интеллектуальных возможностей отдельных субъектов. Для успешного выполнения белком GAP-43 и некоторыми другими белками свойственных им функций необходимо
15. Современные представления о возникновении жизни
15. Современные представления о возникновении жизни Вспомните!Какие химические элементы входят в состав белков и нуклеиновых кислот?Что такое биологические полимеры?Какие организмы называют автотрофами; гетеротрофами?Теория биохимической эволюции. Наибольшее
Эволюционно-генетические проблемы поведения животных и человека[51]
Эволюционно-генетические проблемы поведения животных и человека[51] Известно, что одним из открытий И. П. Павлова явилось обнаружение патологии поведения животных (а также человека) при чрезмерном напряжении высшей нервной деятельности. Большой вклад в исследование
Социальное устройство групп и диета у австралопитековых
Социальное устройство групп и диета у австралопитековых О диете животных можно судить по строению их пищеварительного тракта и особенностям зубной системы. Последнее оказывается исключительно важным для палеоантропологии.Ранние гоминины (австралопитековые) питались