Возникновение и наследование модификаций на примере Metazoon
Возникновение и наследование модификаций на примере Metazoon
Еще один важный теоретический вопрос, который можно рассмотреть на нашем примере с Metazoon, — это вопрос о возникновении адаптивных модификаций. Так называют способность организма более или менее осмысленно (то есть целесообразно) менять свое поведение, строение и онтогенез в зависимости от внешних условий, причем без изменений генома. Простейший пример — мышцы увеличиваются от упражнения.
Модификации — это типичные «благоприобретенные признаки». Синтетическая теория эволюции отрицает возможность наследования модификаций, а ламаркизм основан на признании возможности их наследования. Сам Дарвин в этом отношении был ламаркистом: он считал, что наследование модификаций, возникающих в ходе упражнения органов, вполне возможно.
Вопрос о наследовании приобретенных признаков мы рассмотрим в главе 8, а сейчас займемся другой проблемой, может быть, даже более важной: откуда берется сама адаптивная модификация? Ведь ясно, что способность адекватно изменяться в зависимости от условий — это особый сложный механизм, он не может просто «быть заложен изначально» в свойствах живой материи. Способность к полезным модификациям вырабатывается в ходе эволюции, как и любые другие приспособительные свойства организма. В дальнейшем «модифицированное» состояние может зафиксироваться и стать наследственной нормой. Но самое главное — понять, как возникают новые модификации, ведь именно в этот момент происходит эволюционное новообразование. Многие биологи-теоретики справедливо полагают, что превращение модификации в стойкий наследуемый признак — это акт упрощения, а не усложнения. Действительно, организм, способный к модификации, имеет два или более возможных путей развития, из которых он выбирает один в зависимости от условий. Организм, у которого один из этих путей наследственно закрепился и стал единственно возможным, явно проще исходного.
Посмотрим, каким образом у Metazoon модификация может развиться на основе случайной изменчивости. Допустим, существует Metazoon, у которого из всего разнообразия потенциально возможных жизненных циклов реализуется только два — «двухклеточный» и «четырехклеточный».
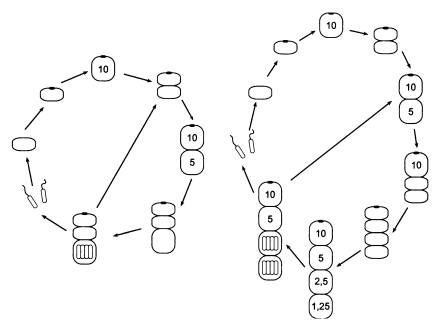
Будем считать, что у этого организма клетки с Px < 4 приступают к мейозу раньше, чем одновременно созревшие клетки с Px > 4 приступают к митозу. Допустим, что это свойство уже зафиксировалось в геноме и не подвержено изменчивости. Это позволит четырехклеточному организму не превращаться чуть что в восьмиклеточный, а нам упростит изложение.
Какой цикл из двух возможных будет избран, изначально определяется чистой случайностью. Все зависит от того, какая из клеток двухклеточного организма поделится первой — передняя или задняя, а это вполне может быть делом случая. Такую ситуацию легко себе представить. Обе клетки находятся на одной стадии зрелости, поэтому то, какая из них начнет делиться чуть раньше, может зависеть от массы случайных факторов.
Теперь представим себе, что в популяции начал действовать отбор. Например, некое хищное существо стало избирательно заглатывать четырехклеточники, не трогая при этом ни двух-, ни одноклеточные стадии.
Следовательно, станут избирательно выживать и размножаться те Metazoon, у которых на двухклеточной стадии передняя клетка делится первой и которые поэтому не достигают четырехклеточной стадии. В этой изменившейся ситуации ни нас, ни Metazoon уже не удовлетворят разговоры о «чистой случайности»! Придется разбираться: от чего же все-таки зависит, какая из двух клеток поделится первой?
В любой живой клетке все процессы связаны в единую сложную сеть. В конечном счете каждое вещество, каждое химическое превращение и каждый регуляторный эффект через более или менее длинную цепочку промежуточных звеньев влияет на все остальные. Большинство этих влияний — слабые, побочные, не имеющие реального значения. Но мы должны помнить, что на любую функцию клетки — в том числе и на скорость митоза — в той или иной степени влияют тысячи белков (каждый из которых может существовать в популяции в виде нескольких разных вариантов — аллелей) и тысячи «малых молекул», поступающих в клетку извне или образующихся в ней, являющихся субстратами, продуктами и промежуточными этапами обмена веществ.
Материалом для отбора являются в первую очередь аллельные варианты генов, кодирующих белки. Жесткий отбор немедленно даст преимущество такому сочетанию этих вариантов, при которых через любую по длине и качеству цепочку взаимодействий сигнал, пусть самый слабый, будет передан из точки А в точку Б регуляторной сети. Здесь «точка А» — это любая из тех особенностей, которые отличают переднюю клетку от задней в двухклеточном организме. «Точка Б» — это сдвиг момента начала митоза (на «чуть раньше» для передней клетки или «чуть позже» для задней).
Учитывая огромную сложность регуляторной сети в любой клетке, практически невозможно предсказать, за какой из множества возможных путей передачи сигнала из А в Б зацепится отбор. Это должна быть такая цепочка, реализация (усиление) которой не принесет заметного ущерба жизнеспособности организма. Цепочка может оказаться как короткой, так и длинной. Она может оказаться неожиданной в том смысле, что будут задействованы белки, не имеющие прямого отношения ни к митозу, ни к отличиям передней клетки от задней. Но именно через них пройдет путь передачи сигнала, потому что были выполнены два условия:
1. именно эта цепочка первой «подвернулась под руку» естественному отбору. Или, говоря строже, среди имеющихся в популяции генетических вариантов оказались такие, в которых именно по этой цепочке из точку А в точку Б уже передавался слабенький «сигнал», в результате чего митоз передней клетки двуклеточного организма чуть ускорялся (или чуть замедлялся митоз задней клетки);
2. «усиление» этой регуляторной цепочки под действием отбора не вызвало вредных побочных эффектов.
В нашем случае известно, что передняя клетка отличается от задней вдвое большей концентрацией вещества X и что уже существует регуляторная система, определяющая выбор способа деления в зависимости от концентрации X (следовательно, есть «датчики», регистрирующие эту концентрацию). С большой вероятностью эти готовые блоки будут задействованы в формировании новой регуляторной связи. Эволюция создает новые структуры из того, что под рукой.
Поначалу некоторым регуляторным белкам, возможно, придется взять на себя новую функцию вдобавок к старой; в дальнейшем гены этих белков могут дуплицироваться (удвоиться), и произойдет разделение функций.
Очень важно понять, что для формирования принципиально нового (чуть ли не любого!) регуляторного эффекта в организме, скорее всего, уже есть преадаптации — своего рода «заготовки». Это следует из самого факта взаимосвязанности всех клеточных процессов.
Но мы пока еще не пришли к настоящей адаптивной модификации. Мы рассмотрели только процесс наследственного закрепления одного из двух «случайных» вариантов онтогенеза. Такой процесс соответствует представлениям о стабилизирующем отборе[56] (произошла «стабилизация» онтогенеза) и ведет к снижению изменчивости. У нас был организм с двумя возможными вариантами развития, один из которых выбирался случайным образом. Теперь мы пришли к организму, у которого всегда выбирается один и тот же вариант (двухклеточный). Этот организм отличается от исходного тем, что у него возникла новая регуляторная связь, новый «оператор условного перехода» в программе поведения клетки. Примерно такой: «если я — передняя из двух клеток, то я делюсь быстрее».
Стал ли организм сложнее исходного, возникла ли новая сложность? На мой взгляд, да. Мне представляется, что именно формирование новой регуляторной связи в программе поведения клетки и является базовым элементарным актом усложнения организма. Однако если смотреть не на программу, а на результат ее выполнения, то есть на сам организм, то усложнение в данном случае выглядит неочевидным. Можно даже подумать, что возможности организма сузились, ведь из двух имевшихся вариантов развития остался только один.
Чтобы усложнение стало совсем уж бесспорным, мы должны сделать еще один шаг и посмотреть, каким образом организм, родившийся в нашем предыдущем мысленном эксперименте, вырабатывает способность к настоящей адаптивной модификации. Оказывается, для этого достаточно создать еще одну регуляторную связь.
Допустим, тот хищник, который избирательно поедает четырехклеточные организмы, активен только при определенных условиях среды — например, при затяжных дождях (назовем эти условия У1). При иных условиях — например, при ясной погоде (У2) — хищник не опасен для Metazoon. Допустим, он откочевывает в другую часть водоема, где Metazoon не живет, и там охотится на кого-то другого. И в условиях У2 преимущество получают четырехклеточные, а не двухклеточные особи Metazoon.
В такой ситуации создаются предпосылки для возникновения новой регуляторной связи, которая будет передавать сигнал от какого-нибудь «датчика внешних условий» к той системе регуляции скорости митоза в зависимости от положения клетки в двухклеточном организме, которая возникла до этого. «Входными данными» для этого нового регуляторного контура будут служить внешние условия (У1 или У2). Скорее всего, эти условия хоть как-нибудь, хоть немного, но будут отражаться на внутренней среде клетки, поэтому отбору будет за что зацепиться. В качестве одного из вариантов можно предположить, что сигналом о наступлении условий У2 будет служить отсутствие запаха хищника.
На выходе новый регуляторный контур должен взаимодействовать с одним из элементов старого контура. В случае наступления условий У2 новый контур будет просто-напросто блокировать старый, отключать его. Программа поведения клетки будет теперь выглядеть так:
Если условия = У1, то
если я — передняя из двух клеток, то
я делюсь быстрее.
Таким образом, теперь система ускорения митоза в передней клетке будет срабатывать только в том случае, если условия = У1 (или, что то же самое, если условия ? У2), то есть в случае присутствия хищника. Если же хищника нет, ускорение митоза в передней клетке блокируется и восстанавливается исходная ситуация случайного выбора одного из двух вариантов развития.
Это уже самая настоящая способность к адаптивной модификации, хотя и не доведенная до полного совершенства. Если хищник присутствует, все особи развиваются по двухклеточному пути; если хищника нет, то часть особей развивается по двух-, а часть — по четырехклеточному пути. Думаю, что читатели, особенно те из них, кто немного знаком с программированием, легко смогут сами придумать, как довести эту систему до совершенства, то есть заставить организмы в условиях У1 (когда хищника нет) всегда выбирать четырехклеточный путь развития.
Похоже на то, что, добавляя понемножку новые «если» в программу поведения клетки, можно получить из нашего Metazoon организм чуть ли не любой степени сложности. Мне кажется, на этой основе можно было бы сделать занятную компьютерную игру.
Теперь уже никак нельзя сомневаться в том, что на наших глазах система стала сложнее — произошло рождение новой сложности. По-видимому, именно таким способом — путем создания дополнительных регуляторных контуров, новых «операторов условного перехода» в программе поведения клетки — усложнялось в ходе эволюции строение многоклеточных организмов, в особенности животных, о которых пойдет речь в следующей главе.
Перейти от достигнутого нашим Metazoon высокоразвитого состояния к одному фиксированному жизненному циклу очень просто. Для этого не надо создавать ничего нового, напротив, нужно упростить систему. Это называют «генокопированием модификаций». В результате «генокопирования» один из вариантов развития будет жестко закреплен в геноме. Он начнет реализовываться всегда, вне зависимости от условий. Представим себе, что условия У2 (хищника нет) перестали возникать, среда стала более стабильной: всегда существуют только условия У1 (хищник есть). Четырехклеточный вариант жизненного цикла становится ненужным. Регуляторный контур, обеспечивающий отключение ускорения митоза передней клетки в отсутствие хищника, больше не задействуется и просто отмирает под грузом случайных мутаций. Мы вернулись к тому же состоянию, которое существовало после первого этапа нашего мысленного эксперимента (когда первая регуляторная связь уже сформировалась, а вторая — еще нет).
Регуляторную цепочку можно представить себе как последовательность переключателей, где изменение регистра первого переключателя приводит к переключению следующего, и т. д. Если в течение многих поколений все переключатели остаются в одном и том же положении, они «ржавеют» и застопориваются в нем навсегда, а затем могут и вовсе «рассосаться». Случайные мутации и отбор — точнее, в данном случае, его отсутствие, поскольку особи с «ржавыми» переключателями перестают выбраковываться — прекрасно объясняют этот процесс упрощения. В конце концов остается последний «ржавый», навсегда включенный переключатель, вплотную примыкающий к регулируемому процессу. В нашем случае это будет выражаться в том, что в определенном месте регуляторной цепочки, идущей от «порядкового номера клетки» к митозу — а именно там, где раньше эта цепочка сливалась с другой, идущей от «внешних условий», — будет стоять «ржавый переключатель», якобы показывающий, что условия = У1
Вот у нас и унаследовалась модификация. Правда, не сразу («по Ламарку»), а за ряд поколений («по Дарвину»). Как видим, ничего хитрого в механизме наследования модификаций нет, одно сплошное упрощение.
—————
Почему эволюция иногда сохраняет «ненужные» регуляторные контуры? Любопытно, что в некоторых случаях регуляторные контуры могут в течение миллионов лет сохранять способность адекватно реагировать на изменения положения тех «переключателей», которые давным-давно «заржавели» или вовсе исчезли. Современные методы иногда позволяют «воскресить» отмерший переключатель, и тогда появляется возможность наглядно убедиться в том, что система по-прежнему в рабочем состоянии. Таким способом, например, удалось вырастить зачатки зубов у куриного эмбриона, воздействовав на ткани челюсти соответствующими регуляторными веществами, взятыми у других животных. Предки курицы утратили зубы десятки миллионов лет назад, но многие компоненты регуляторного контура, необходимого для роста зубов, сохранились! Почему они не рассыпались под грузом случайных мутаций? Дело в том, что у животных важнейшие блоки регуляторных каскадов являются многофункциональными, то есть входят в разных комбинациях в состав множества регуляторных контуров. Поэтому отбор оберегает эти блоки, отсеивая мутации, ведущие к их повреждению. Еще один пример «воскрешения отмершего переключателя» мы рассмотрим в главе «Животные» (см. сюжет «Разгадан механизм регенерации конечностей»): активировав один-единственный регуляторный ген, ученым удалось вернуть куриному эмбриону способность к регенерации конечности, давным-давно утраченную предками птиц.
—————
Что же касается нашего Metazoon со способностью к адаптивной модификации, то он — существо весьма перспективное.
Возникшие у него новые регуляторные каскады открывают великолепные возможности для дальнейшей эволюции. Сначала новшества могут появляться за счет мелких мутаций в регуляторных генах, чуть меняющих реакцию белков на различные сигналы, или в результате попадания организма в непредвиденную ситуацию, на которую не были рассчитаны регуляторные связи и в которых они начнут работать немного иначе. В дальнейшем эти новые «случайные отклонения» могут закрепляться или как основная норма, или как новая адаптивная модификация.
—————
Выведена гусеница, меняющая цвет при нагревании. Недавно возможность появления новой адаптивной модификации (полифенизма) под действием отбора была продемонстрирована экспериментально. Американские ученые путем искусственного отбора вывели гусениц, цвет которых дискретно меняется в зависимости от температуры: в тепле развиваются зеленые гусеницы, на холоде — черные.
—————
Норма реакции и полифенизм. Многие далекие от генетики люди полагают, что в генах «записано» строение организма (генотип определяет фенотип). Это не совсем так. В действительности генотип определяет не фенотип как таковой, а норму реакции — определенный спектр возможностей развития. Какая из этих возможностей будет реализована, зависит уже не от генов, а от условий, в которых будет происходить развитие организма.
Обычно эти предусмотренные генотипом допустимые варианты фенотипа образуют непрерывный, плавный ряд, но иногда они бывают дискретны. В таком случае говорят о явлении полифенизма. Это значит, что при одном и том же генотипе в зависимости от условий будет реализован один из нескольких дискретных вариантов фенотипа. Например, из муравьиного яйца с одним и тем же генотипом могут развиться и рабочий муравей, и крылатая самка — в зависимости от того, как будут кормить личинку. Другие примеры полифенизма — одиночная и стайная формы перелетной саранчи, бескрылая и крылатая формы тлей.
Полифенизм — одна из форм адаптации к переменчивым условиям среды. Известно, что пороговые значения внешних факторов, вызывающие «переключение» развития на альтернативный путь, могут меняться под воздействием отбора. Однако сам механизм этого переключения пока мало изучен.
—————
В качестве объекта выбрали бабочку Manduca sexta. У этого вида полифенизма нет, однако он есть у близкородственного вида М. quinquemaculata, у которого при 20 °C развиваются черные гусеницы, а при повышении температуры до 28 °C — зеленые.
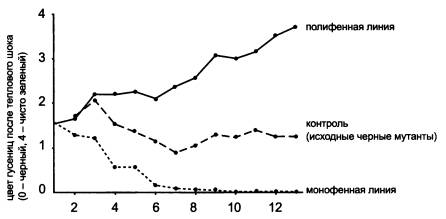
Цвет гусениц после теплового шока в ходе отбора становился все более зеленым в полифенной линии и все более черным — в монофенной.
У Manduca sexta вне зависимости от температуры гусеницы зеленые, однако встречаются и мутанты — черные гусеницы. Но это не полифенизм, а обыкновенная наследственная изменчивость (полиморфизм), поскольку цвет гусеницы определяется генотипом, а не внешними условиями.
Мутация black («черная окраска») приводит к уменьшению выработки так называемого ювенильного гормона, что в свою очередь ведет к меланизации (почернению) покровов. Нормальные (не мутантные) зеленые гусеницы не меняют свой цвет ни при каких обстоятельствах, однако ученые обнаружили, что окраска черных мутантов менее стабильна. Если подвергнуть гусеницу-мутанта в определенном возрасте шестичасовому тепловому шоку (нагреванию до 30–33 °C), то после очередной линьки гусеницы получаются разные — черные, с прозеленью или почти нормального зеленого цвета.
Ухватившись за это обстоятельство, ученые приступили к искусственному отбору гусениц с мутацией black. В одной линии отбирались гусеницы, реагирующие на тепловой шок минимально (т. е. остающиеся черными или почти черными). В другой линии, наоборот, отбирались те особи, которые реагировали на перегрев максимальным позеленением. Уже в седьмом поколении в первой линии была полностью утрачена способность реагировать на тепловой шок изменением окраски (полный монофенизм).
Во второй линии за 13 поколений возник настоящий полифенизм. Все гусеницы в этой линии теперь становились ярко-зелеными при постоянной температуре 28 °C. Теплового шока, нагревания до 30–33 °C, уже не требовалось. При более низких температурах гусеницы оставались черными.
Чтобы проверить, связано ли изменение окраски у полифенных гусениц с ювенильным гормоном, ученые провели следующий жестокий эксперимент: туго перевязали гусениц поперек брюшка, чтобы гормон, синтезирующийся в голове и груди, не мог попасть в заднюю часть тела. Гусеницы стоически перенесли издевательство и доказали правильность предположений своих мучителей, позеленев спереди и оставшись черными сзади. Гусеницы из монофенной линии, естественно, оставались черными хоть с перевязками, хоть без.
Результаты продемонстрировали тот путь, который проделывает случайная мутация (такая как мутация black) под действием отбора до настоящего полифенизма, и что изменения гормональной регуляции могут играть важную роль в подобных эволюционных преобразованиях.
Более общий эволюционный вывод, который можно сделать на основании этой работы, состоит в том, что генотип может содержать в себе в скрытом виде разнообразные новые возможности (варианты развития), до поры до времени не проявляющиеся, замаскированные различными регуляторными механизмами — например, той же гормональной регуляцией. Но в экстраординарных условиях, в результате мутации или стресса, регуляторные механизмы могут дать сбой, и тогда скрытая изменчивость станет явной и может стать основой для развития новых полезных адаптаций.
(Источник: Suzuki Y., Nijhout H. F. Evolution of a Polyphenism by Genetic Accommodation // Science. 2006. V. 311. P. 650–652.)
—————
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКДанный текст является ознакомительным фрагментом.
Читайте также
«Направленные мутации» и наследование соматических мутаций
«Направленные мутации» и наследование соматических мутаций Мы описали протекающий в В-лимфоцитах процесс обратной связи V(D)J-генов, основанный на склонной к ошибкам обратной транскрипции. Он составляет основу определяемого антигеном мутирования генов антител. Все
Наследование соматических мутаций
Наследование соматических мутаций Итак, мы можем спросить, есть ли польза от антигензависимых соматических событий (мутаций и отбора) для ДНК-последовательностей V-генов зародышевой линии следующих поколений? В предьщущей главе мы уже рассказали, как много появляется
Наследование мозолистых утолщений
Наследование мозолистых утолщений Сейчас мы рассмотрим некоторые явления наследования анатомических признаков в экспериментах, проведенных природой. Некоторые из них детально описаны Джоунзом в его книге 1943 г. Поскольку животные «сами приноравливают себя к условиям
Эпигенетическое наследование
Эпигенетическое наследование Нашу экспериментальную и теоретическую работу по «ретрогенетике» V-генов иммунной системы можно рассматривать как пример жесткого наследования, затрагивающего генетическую информацию, воплощенную в последовательности оснований ДНК.
Наследование, сцепленное с полом
Наследование, сцепленное с полом В предыдущих двух случаях, когда мы рассматривали наследование врожденных аномалий по доминантному и по рецессивному типу, предполагалось, что мутации затрагивают аутосомы (греч. autos — сам и soma — тело), то есть находятся не в половых
Эпигенетическое наследование
Эпигенетическое наследование Недавно открыто еще несколько молекулярных механизмов, которые в принципе могут служить для передачи по наследству приобретенных признаков. Эти механизмы не связаны напрямую с изменениями самого «текста», записанного в структуре молекул
4.5. Генетика пола и сцепленное с полом наследование
4.5. Генетика пола и сцепленное с полом наследование Генетический механизм определения пола в природе обусловлен генами, локализованными на особых половых хромосомах, имеющихся в кариотипе. Пол, у которого в кариотипе одинаковые половые хромосомы, называется
Ничто в природе слепо не стремится к совершенству – на примере… орхидей
Ничто в природе слепо не стремится к совершенству – на примере… орхидей В природе, следовательно, не существует слепого стремления к прогрессу. Ежедневно зарождается множество организмов с изменчивыми признаками.Те из них, которые хорошо приспособляются к внешним
Возникновение сегментов
Возникновение сегментов Сегментация зародыша дрозофилы в своей основе выглядит как ряд латеральных впячиваний зародышевой полоски, образующихся почти одновременно на стадии гаструляции. Несмотря на кажущийся мозаичный характер этого процесса, можно показать, что
Глава 7. Наследование формы
Глава 7. Наследование формы 7.1. Генетика и наследственность Наследственные различия между организмами, одинаковыми в других отношениях, зависят от генетических различий; эти последние зависят от различий в структуре ДНК или в ее расположении в хромосомах, а эти различия
7.7. Наследование приобретенных признаков
7.7. Наследование приобретенных признаков Влияние предшествующих организмов на последующие подобные организмы путем морфического резонанса может приводить к эффектам, которые невозможно было бы предположить в случае, если наследственность зависит только от передачи
Глава 11. Наследование и эволюция поведения
Глава 11. Наследование и эволюция поведения 11.1. Наследование поведения Согласно гипотезе формативной причинности, наследование поведения зависит от генетической наследственности, а также от морфогенетических полей, которые контролируют развитие нервной системы и
11.1. Наследование поведения
11.1. Наследование поведения Согласно гипотезе формативной причинности, наследование поведения зависит от генетической наследственности, а также от морфогенетических полей, которые контролируют развитие нервной системы и всего животного в целом, а также от моторных
Глава 4. ЭВОЛЮЦИЯ ЕСТЕСТВЕННЫХ ТЕХНОЛОГИЙ НА ПРИМЕРЕ ЭКЗОТРОФИИ
Глава 4. ЭВОЛЮЦИЯ ЕСТЕСТВЕННЫХ ТЕХНОЛОГИЙ НА ПРИМЕРЕ ЭКЗОТРОФИИ Экзотрофия, и в том числе пищеварение, была охарактеризована как некоторая естественная технология. В этой главе такая технология будет рассмотрена в эволюционном аспекте.Было бы привлекательно думать, что
9.3. Возникновение клеток
9.3. Возникновение клеток Предполагается, что этапом возникновения жизни на Земле следует считать период, когда сформировались простейшие клеточные системы, ставшие элементарной ячейкой живого. Сведения, касающиеся этой проблемы, освещены в обзорах, посвященных