2.3. Основные типы пищеварения
2.3. Основные типы пищеварения
По механизму действия ферментов на субстраты, по механизмам их поступления к месту функционирования, по взаимоотношениям пищеварительных процессов и клеточной границы и, наконец, по отношению к процессам транспорта можно выделить три основных типа пищеварения: внеклеточное дистантное, внутриклеточное и мембранное (рис. 4).
2.3.1. Внеклеточное дистантное пищеварение
Этот тип пищеварения характеризуется тем, что синтезированные секреторными клетками ферменты выделяются во внеклеточную среду, где реализуется их гидролитический эффект. Этот тип пищеварения является основным у организмов, стоящих на более высоком этапе эволюционного развития, чем плоские черви, и преобладает у кольчатых червей, ракообразных, насекомых, головоногих, оболочников и хордовых. Он особенно развит у высокоорганизованных животных и человека, у которых обеспечивает начальное переваривание пищевых веществ. У этих организмов секреторные клетки расположены достаточно далеко от полостей, где реализуется действие ферментов, поэтому внеклеточное пищеварение определяется как дистантное. Дистантное пищеварение, происходящее в специальных полостях, обозначается как полостное. Дистантное пищеварение может происходить и за пределами организма, продуцирующего ферменты. Так, насекомые вводят пищеварительные ферменты в обездвиженную добычу, а бектерии выделяют разнообразные ферменты в культуральную среду.
Так как секретируемые в составе пищеварительных соков ферменты растворены в водной фазе, их пространственная организация невозможна или весьма ограничена. Если в растворе действует несколько ферментов, то их распределение имеет вероятностный характер. Растворенные ферменты эффективны при расщеплении как поверхностно, так и глубоко локализованных пептидных, глюкозидных, эфирных и других связей благодаря любой ориентации активного центра фермента относительно атакуемых молекул субстрата.
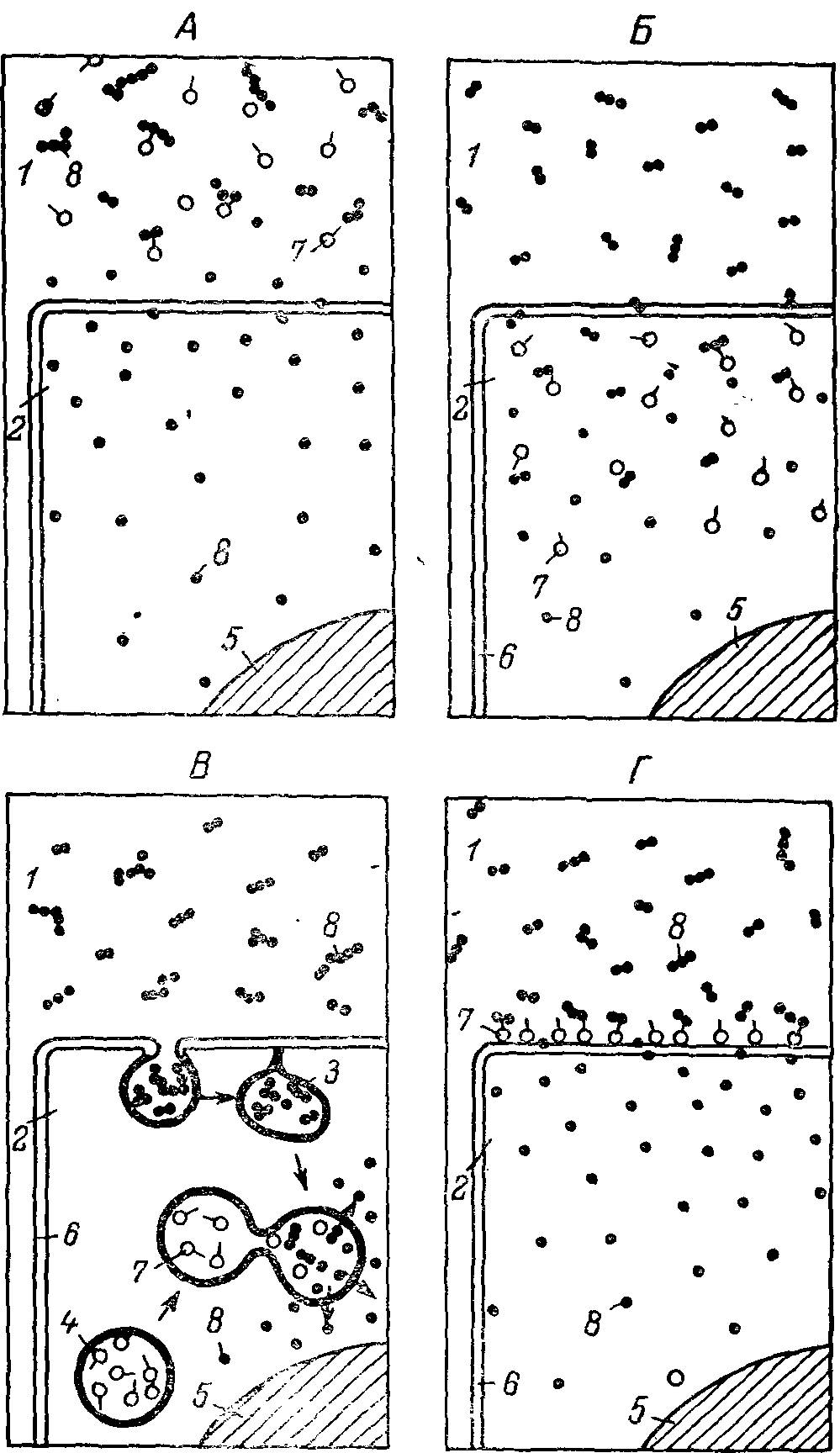
Рис. 4. Основные типы пищеварения.
А — внеклеточное дистантное пищеварение; Б — внутриклеточное цитоплазматическое пищеварение; В — внутриклеточное вакуолярное, или вне-плазматическое, пищеварение, связанное с эндоцитоэом (фаго- и пиноцитозом); Г — мембранное пищеварение. 1 — внеклеточная среда; 2 — внутриклеточная среда; 3 — внутриклеточная пищеварительная вакуоль; 4 — лизосома; 5 — ядро; 6 — мембрана; 7 — ферменты; 8 — субстраты и продукты их гидролиза.
Вместе с тем целостное пищеварение не обеспечивает эффективного перехода от гидролиза к транспорту, так как резорбирующая мембрана и освобождающиеся в процессе гидролиза мономеры разделены значительным расстоянием и требуется определенное время, чтобы мономеры достигли поверхности всасывающей клетки. Существует определенная вероятность перехвата продуктов гидролиза (прежде чем они достигнут резорбирующей поверхности) бактериями, населяющими полость тонкой кишки высших организмов.
2.3.2. Внутриклеточное пищеварение
Этим термином обозначаются случаи, когда не-расщепленные или частично расщепленные пищевые вещества проникают внутрь клетки, где подвергаются гидролизу ферментами цитоплазмы, не выделяемыми за пределы клетки. Внутриклеточное пищеварение распространено у простейших и наиболее примитивных многоклеточных организмов, например у губок и плоских червей. Как дополнительный механизм гидролиза пищевых веществ оно встречается у немертин, иглокожих, некоторых кольчатых червей и многих моллюсков. У высших позвоночных животных и человека оно выполняет главным образом защитные функции, например фагоцитоз.
Различаются два типа внутриклеточного пищеварения. Первый связан с транспортом небольших молекул через клеточные мембраны и последующим перевариванием ферментами цитоплазмы. Внутриклеточное пищеварение может также происходить в специальных внутриклеточных полостях — пищеварительных вакуолях, присутствующих постоянно или образующихся при фагоцитозе и пиноцитозе и исчезающих после расщепления захваченной пищи. Второй тип пищеварения в большинстве случаев связан с участием лизосом, которые содержат широкий набор гидролитических ферментов (фосфатаз, протеаз, глюкозидаз, липаз и др.) с оптимумом действия в кислой среде (pH 3.5—5.5). Пищевые структуры или пищевые растворы в околоклеточной среде вызывают впячивания плазматической мембраны, которые затем отшнуровываются и погружаются в цитоплазму, образуя пиноцитозные и фагоцитозные вакуоли. Соединяясь с последними, лизосомы образуют фагосомы, где происходит контакт ферментов с соответствующими субстратами. Образовавшиеся продукты гидролиза всасываются через мембраны фагосом. После завершения пищеварительного цикла остатки фагосом выбрасываются за пределы клетки путем экзоцитоза. Лизосомы играют также важную роль в расщеплении собственных структур клетки, которые используются в качестве пищевого материала либо данной клеткой, либо за ее пределами.
По своим механизмам внутриклеточное пищеварение может быть рассмотрено как сочетание микрополостного и мембранного гидролиза в пределах клетки. Действительно, при внутриклеточном пищеварении ферменты могут оказывать свой гидролитический эффект в цитоплазме клетки или в фагосоме, т.е. в среде, что свойственно полостному пищеварению, а также на внутренней поверхности фагосомной мембраны, что свойственно мембранному пищеварению.
Внутриклеточное пищеварение лимитировано проницаемостью мембраны и процессами эпдоцитоза, которые характеризуются небольшой скоростью и, по-видимому, не могут играть существенной роли в обеспечении пищевых потребностей высших организмов.
Тем не менее они могут способствовать проникновению в клетку некоторых уникальных веществ, в частности иммуноглобулинов.
Эндоцитозам приписывается важная роль в ассимиляции пищевых веществ в период раннего постнатального развития.
2.3.3. Мембранное (пристеночное, контактное) пищеварение
Мембранное пищеварение, открытое в конце 50-х гг., пространственно занимает промежуточное положение между внеклеточным и внутриклеточным и осуществляется ферментами, локализованными на структурах клеточной мембраны и ее дериватов (у высших животных и человека — на апикальной поверхности кишечных клеток). Активные центры ферментов ориентированы определенным образом по отношению к мембране и водной фазе. Свободная ориентация каталитических центров ферментов по отношению к субстратам невозможна. Глубоко расположенные связи, по-видимому, недоступны действию ферментов, осуществляющих мембранное пищеварение. Этим оно существенно отличается от полостного и внутриклеточного типов, если последнее происходит в фагосомах.
Мембранное пищеварение осуществляется как адсорбированными из полости тонкой кишки ферментами (преимущественно панкреатического происхождения), так и собственно кишечными, или мембранными, синтезированными в кишечных клетках и встроенными в состав их апикальной липопротеиновой мембраны (табл. 1). Ферменты, адсорбированные на структурах кишечной слизистой (главным образом в гликокаликсном пространстве), реализуют в основном промежуточные этапы гидролиза всех основных видов пищевых веществ. Собственно кишечные ферменты осуществляют преимущественно заключительные этапы расщепления пищевых биополимеров. По-видимому, адсорбированные ферменты связаны в основном со структурами гликокаликса (рис. 5), а собственно кишечные встроены в структуры плазматической мембраны кишечных клеток. Тем не менее на поверхности липопротеиновой мембраны могут адсорбироваться ферменты, поступающие в полость тонкой кишки с панкреатическим соком, а собственно кишечные ферменты, по крайней мере частично, могут включаться в гликокаликс.
Основные ферменты, реализующие мембранное пищеварение в тонкой кишке млекопитающих
Происхождение фермента Фермент КФ Адсорбированные панкреатические ферменты Амилаза 3.2.1.1 Липаза 3.1.1.3 Трипсин 3.4.21.4 Химотрипсин 3.4.21.1 Карбоксипептидаза А 3.4.12.2 Карбоксипептидаза В 3.4.12.3 Эластаза 3.4.21.11 Рибонуклеазэ 3.1.4.22 Собственно кишечные ферменты Мальтаза 3.2.1.20 Сахараза 3.2.1.48 Изомальтаза 3.2.1.10 Гамма-амилаза 3.2.1.3 Лактаза 3.2.1.23 Трегаяаза 3.2.1.28 Щелочная фосфатаза 3.1.3.1 Моноглицеридлипаза 3.1.1.23 Пептидазы 3.4.11 - 15 Аминопептидаза * 3.4.11.2 Дипептидиламинопепти- даза 3.4.14.1 Карбоксипептидаза 3.4.12.4 Энтерслептидаза 3.4.21.9 Гамма-глутамилтранспептидаза 2.3.2.2 Холестеролэстераза 3.1.1.13* Аминопептидаза М, аминопешчдаза N, аланинаминопептидаза.
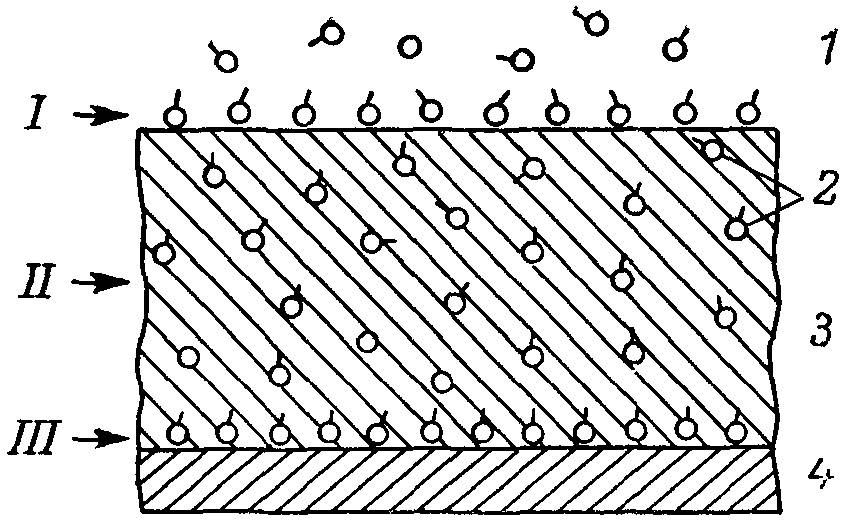
Рис. 5. Упрощенная схема распределения адсорбированных ферментов на поверхности гликокаликса (I), в гликокаликсном пространстве (II) и на липопротеиновой мембране (III) кишечной клетки.
1 — полость тонкой кишки; 2 — ферменты;3 — гликокаликс; 4 — мембрана.
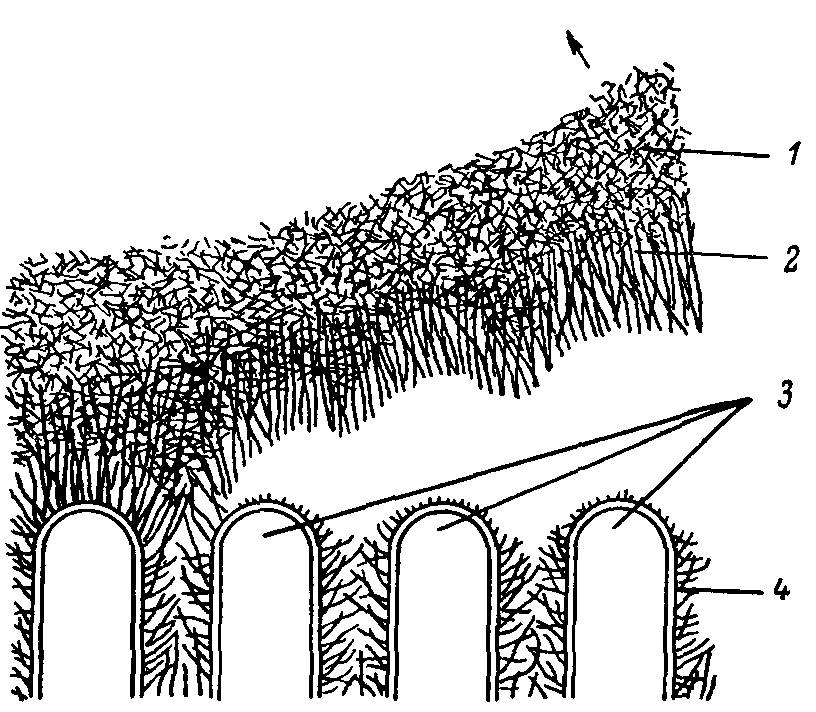
Рис. 6. Схема отделения апикального гликокаликса от липопротеииовой мембраны кишечной клетки.
1 — агаровая реплика; 2 — апикальный гликокаликс; 3 — микровор-синки; 4 — латеральный гликокаликс.
Рис. 7. Щеточная кайма кишечной клетки крысы.
А— продольный разрез апикальной зоны интактной клетки; виден гликокаликс на внешней (апикальной) и боковой (латеральной) поверхностях микроворсинок. В — продольный разрез апикальной зоны клетки после отделения агаровой реплики; гликокаликс на внешней поверхности микроворсинок отсутствует, видны неповрежденная липопротеиновая мембрана клетки и латеральный гликокаликс. 80 000х.
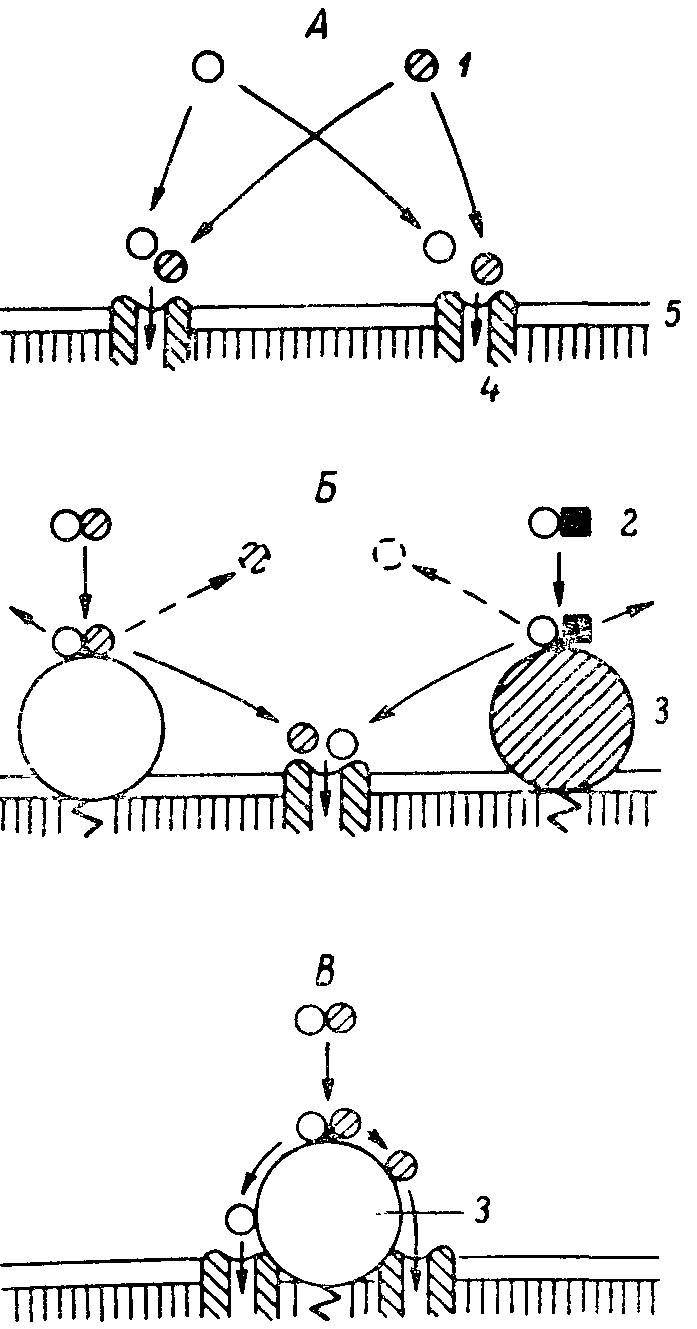
Рис. 8. Роль ферментно-транспортных комплексов в предотвращении конкуренции между мономерами на стадии всасывания.
А — конкуренция между мономерами за общий вход в транспортную систему; Б — конкуренция между финальными продуктами гидролиза за общий вход в транспортную систему; В — ферментно-транспортный комплекс: передача конечных продуктов гидролиза с фермента на вход в транспортную систему (без конкуренции). 1 — мономер; 2 — димер; 3 — фермент; 4 — транспортная система; 5 — мембрана.
Несколько лет назад нами впервые препаративно отделен апикальный гликокаликс от плазматической мембраны кишечных клеток крыс без повреждения этой мембраны (рис. 6, 7). Было обнаружено, что в апикальном гликокаликсе, отделяющем мембрану от внеклеточной среды, сосредоточено около 60% панкреатической амилазы, более 80% трипсина и около 20% химотрипсина, адсорбированных на структурах кишечной слизистой оболочки. Следовательно, примерно 40% амилазы, 20% трипсина и 80% химотрипсина, адсорбированных на этой оболочке, могут быть локализованы в латеральном гликокаликсе, т.е. в межмикрозорсинчатом пространстве, а также, возможно, частично на липопротеиновой мембране. В этих же экспериментах продемонстрировано, что такие собственно кишечные ферменты, как сахараза, гамма-амилаза, ди- и трипептидазы, связаны преимущественно с липопротеиновой мембраной. Однако щелочная фосфатаза, рассматриваемая как трансмембранный интегральный фермент, присутствует в довольно значительных количествах (до 20%) в апикальном гликокаликсе. (Сходные данные получены па курах). Кроме того, выявлено, что в апикальном гликокаликсе содержится 3.6% лейцинариламидазы, менее 2% пролилглициндипептидазы и менее 1% глицилпролинди-пептидазы.
Благодаря локализации кишечных ферментов на липопротеиновой мембране в непосредственной близости от транспортных систем мембранное пищеварение обеспечивает сопряжение конечных этапов переваривания и начальных этапов всасывания. Это достигается в результате специальной организации пищеварительных и транспортных функций клеточной мембраны в виде своеобразного пищеварительно-транспортного конвейера, способствующего передаче конечных продуктов гидролиза с фермента на вход в транспортную систему и предотвращению конкуренции между ними за обладание входом в последнюю (рис. 8). Иными словами, мембранный фермент и транспортная система образуют олигомерный комплекс, между частями которого существуют кооперативные и аллостерические взаимодействия (рис. 9, 10).
Мембранное пищеварение наблюдается у организмов на всех уровнях эволюционного развития, т.е. является универсальным механизмом. Оно обнаружено у всех млекопитающих, включая человека, у птиц, рыб, амфибий, круглоротых, а также у беспозвоночных животных, в том числе у насекомых, ракообразных, моллюсков, у различных паразитирующих форм. Существуют данные о наличии мембранного пищеварения у дрожжей, бактерий и в корнях растений.
Для правильной оценки мембранного пищеварения существенны следующие факторы.
1. Ферменты, реализующие этот процесс, относятся к структурированным. В связи с этим возможна пространственная организация как ферментных, так и транспортных систем, объединяющих заключительные этапы переваривания и начальные этапы всасывания.
2. Структурирование ферментов приводит к изменению их свойств. Так, отделение ферментов от мембраны меняет их каталитические и регуляторные характеристики.
3. Зона мембранного пищеварения обладает особыми физико-химическими свойствами, в частности такими как pH, концентрация органических и неорганических ионов, неперемешиваемый слой жидкости.
4. Мембранное пищеварение осуществляется в стерильной зоне, недоступной для бактерий, что предотвращает поглощение последними легко усвояемых низкомолекулярных пищевых веществ.
5. Благодаря ферментному аппарату гликокаликсный слой превращается в высокоспецифический фильтр. Через этот слой проникают те вещества, для которых на поверхности и внутри гликокаликсного пространства имеются адекватные ферменты, но не проникают другие вещества с такими же размерами молекул.
6. Для понимания эффективности мембранного пищеварения и трансмембранной проницаемости важны сведения о сократительной функции щеточной каймы, регулируемой кальцием. Показано, что микрофиламенты микроворсинок, выполняющие эти функции, содержат актин и миозин и связаны с апикальной мембраной кишечных клеток.
7. Следует учитывать не только процессы синтеза кишечных ферментов и их включения в состав апикальной мембраны кишечных клеток, по и скорость их деградации. (Более подробно см. гл. 5).
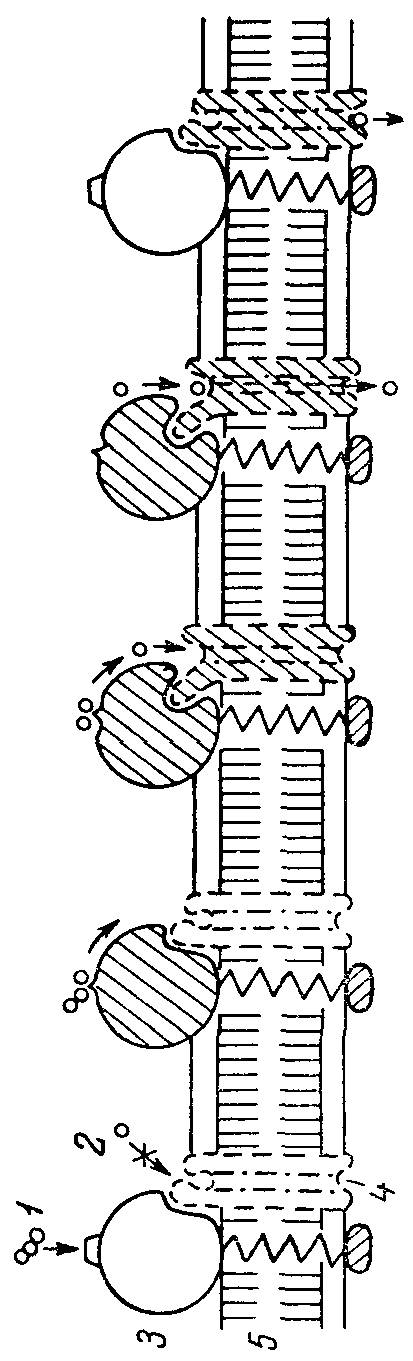
Рис 9. Схема последовательных конформационных взаимодействий и транспортной частей комплекса.
1 - субстрат; 2 - продукт; 3 - трансмембранный фермент; транспортная система; 5 - мембрана
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Тема 9. ВОЗРАСТНЫЕ ОСОБЕННОСТИ ПИЩЕВАРЕНИЯ
Тема 9. ВОЗРАСТНЫЕ ОСОБЕННОСТИ ПИЩЕВАРЕНИЯ 9.1. Строение пищеварительного канала Пищеварительный канал состоит из системы органов, которые производят механическую и химическую обработку пищи и ее всасывание. У человека пищеварительный канал имеет вид трубки длиной
9.2. Процесс пищеварения
9.2. Процесс пищеварения Особенности пищеварения в желудке. Желудок – наиболее расширенная часть пищеварительной системы. Он имеет вид изогнутого мешка, вмещающего до 2 л пищи.Расположен желудок в брюшной полости асимметрично: большая его часть находится слева, а меньшая
Разгадка тайны долгожительства кроется в органах пищеварения
Разгадка тайны долгожительства кроется в органах пищеварения Напрасно стали бы мы искать какой-нибудь точки опоры для объяснения значительного укорачивания жизни млекопитающих сравнительно с птицами или так называемыми «холоднокровными» позвоночными в устройстве их
Глава VIII Система пищеварения
Глава VIII Система пищеварения Пища и питание Мне всегда нравилось выражение «Мы — то, что мы едим»; это относится к кошкам и собакам в той же степени, что и к человеку. От неполноценной пищи прежде всего страдает система пищеварения; другие органы — печень, поджелудочная
Болезни органов пищеварения Л. В. Панышева
Болезни органов пищеварения Л. В. Панышева Болезни органов пищеварения занимают одно из главных мест среди незаразных заболеваний собак. Особенно часто расстройством пищеварения страдают щенки после отъема от матери. Целый ряд инфекционных заболеваний, таких, как
Исследование органов пищеварения
Исследование органов пищеварения Наблюдая за приемом корма и питья, можно заметить целый ряд отклонений. Частичной потерей аппетита или полной его утратой начинаются многие заболевания. Отсутствие аппетита бывает при лихорадке, инфекционных, инвазионных заболеваниях,
Исследование системы органов пищеварения
Исследование системы органов пищеварения При исследовании системы органов пищеварения фиксируются следующие вопросы:– прием собакой пищи и воды;– состояние губ, языка, ротовой полости (зубов);– наличие рвоты, слюнотечения, кровотечения из прямой кишки;–
Заболевания системы органов пищеварения
Заболевания системы органов пищеварения Стоматит Стоматит – воспаление слизистой ротовой полости. Различают стоматиты катаральные, везикулярные, язвенные, афтозные и флегмонозные. Болеют плотоядные всех возрастов и пород.¦ ЭТИОЛОГИЯПервичные стоматиты возникают как
Типы ритмов
Типы ритмов Золотистые водоросли демонстрируют суточный ритм, хотя их сутки и составляют 24,8 ч. Подобные ритмы называются циркадианными (от латинских слов circa — около и dies — день) или околосуточными. Цикл сна и бодрствования у человека, суточные колебания температуры
Глава 2. ЭКЗОТРОФИЯ. МЕХАНИЗМЫ ПИЩЕВАРЕНИЯ
Глава 2. ЭКЗОТРОФИЯ. МЕХАНИЗМЫ ПИЩЕВАРЕНИЯ Метаболический цикл начинается с поступления веществ, необходимых для выполнения живыми системами их основных функций. Все живые организмы нуждаются в пище, т.е. источниках энергии, строительных и пластических материалах, солях
2.4. Схема переваривания пищи как сочетание трех основных типов пищеварения
2.4. Схема переваривания пищи как сочетание трех основных типов пищеварения После обнаружения мембранного пищеварения классическая схема ассимиляции пищи претерпела существенные изменения. Согласно классическим представлениям, пищевые вещества — нутриенты, способные
2.5. Специализированные механизмы пищеварения
2.5. Специализированные механизмы пищеварения Пищеварительные процессы можно классифицировать не только по механизмам (внеклеточное дистантное, внутриклеточное, мембранное), но и по источникам ферментов. В зависимости от этого критерия можно выделить три типа
4.1. Классические теории эволюций пищеварения
4.1. Классические теории эволюций пищеварения С конца XIX в. ж до последнего времени была принята концепция происхождения и эволюции пищеварения, созданная И. И. Мечниковым в 1880 г., а затем развитая X. Иорданом ж другими исследователями. Согласно этой концепции, пищеварение
4.2. Распространение основных типов пищеварения
4.2. Распространение основных типов пищеварения Прежде всего рассмотрим критерии, на основании которых можно говорить о наличии или отсутствии у животных различных систематических групп внеклеточного, внутриклеточного и мембранного пищеварения. Современные критерии
4.6. Происхождение пищеварения
4.6. Происхождение пищеварения При обсуждении происхождения и эволюции различных форм пищеварения допускается традиционная ошибка. Она заключается в том, что почти всегда обращается внимание на взаимоотношения полостного, внутриклеточного и мембранного типов
3. Система органов пищеварения
3. Система органов пищеварения Организм собаки построен из сложных органических веществ — белков, углеводов, жиров. Важнейшим из них является белок. Кроме этих органических веществ, в организме имеются и неорганические вещества — соли и большое количество воды (от 65 до