5.2. Кишечная клетка
5.2. Кишечная клетка
Схема кишечной клетки представлена на рис. 26. Известно, что численность кишечных клеток составляет 1010, а соматических клеток взрослого человека— 10 15. Следовательно, одна кишечная клетка обеспечивает питание около 100 000 других клеток. Такая эффективность, вероятно, не имеет аналогий в производственных технологиях. На первый взгляд клетка кажется сверхспециализированной и приспособленной к выполнению именно пищеварительных и транспортных функций. В плане нашей задачи особый интерес представляет структурно-функциональная организация апикальной поверхности кишечных клеток, т.е. поверхности, обращенной в полость тонкой кишки.
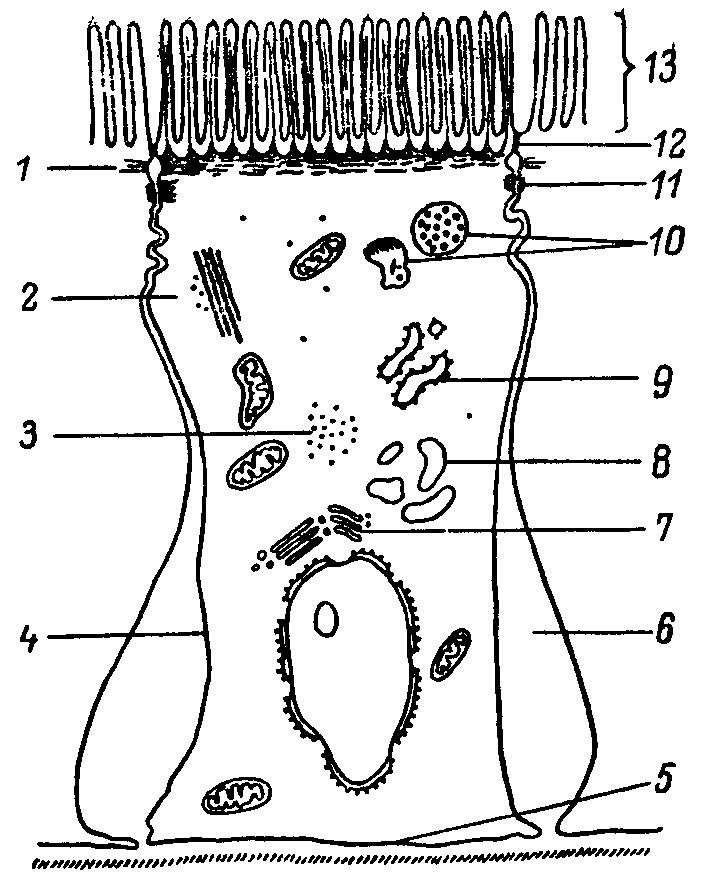
Рис. 26. Схема кишечной клетки.
1 — терминальная сеть; 2 — микротрубочки; 3 — свободные рибосомы; 4 — латеральная мембрана; 5 — базальная мембрана; 6 — межклеточное пространство; 7 — аппарат Гольджи; 8 — гладкий ретикулум; 9 — гранулярный эндоплазматический ретикулум; 10 — лизосомы; 11 — десмосома; 12 — плотный контакт; 13 - микроворсинки.
Щеточная кайма. Характерная особенность кишечной клетки заключается в наличии щеточной каймы, образованной микроворсинками — плазматическими выростами, ограниченными мембраной. Щеточная кайма является универсальной структурой, свойственной клеткам самых различных животных, а также человека. На апикальной поверхности каждой кишечной клетки находится около 3000—4000 микроворсинок; на 1 мм2 поверхности кишечного эпителия приходится до 50— 100 млн. микроворсинок. У человека и других млекопитающих высота микроворсинок в среднем составляет 1 мкм, диаметр в 10 раз меньше (около 0.1 мкм),хотя у низших позвоночных, включая амфибий, микроворсинки могут быть и длиннее. Наименьшее расстояние между микроворсинками — 15—20 нм. Благодаря микроворсинкам поверхность тонкой кишки увеличивается примерно в 40 раз, что способствует включению в мембрану значительного количества различных ферментативно активных и транспортных белков.
Гликокаликс. Внешняя поверхность плазматической мембраны большинства клеток, в том числе кишечных, покрыта гликокаликсом. Последний является компонентом мембраны и образует на апикальной поверхности кишечных клеток слой толщиной до 0.1 мкм. Гликокаликс состоит из мукополисахаридных нитей, или филамент, связанных кальциевыми мостиками и образующими особую сеть. Он обеспечивает не только механическую прочность плазматической мембраны. Благодаря лабильности кальциевых мостиков связи между отдельными филаментами периодически разрушаются, что способствует проникновению относительно крупных молекул в глубину гликокаликса. Так как кислотные остатки гликокаликса имеют отрицательный заряд, то проникающие ионы и диполи будут определенным образом ориентироваться по отношению к гликокаликсу. Таким образом, гликокаликс представляет собой молекулярное сито, сепарирующее молекулы по величине и заряду, имеет отрицательный заряд, характеризуется значительной гидрофильностью и придает процессам переноса пищевых веществ векторный и селективный характер.
Гликокаликс определяет ряд других важнейших функций клеток: «узнавание», связь молекул определенных типов (т.е. специализированные акцепторные и рецепторные функции), иммунологическое дифференцирование, межклеточные взаимодействия и т.д. По-видимому, он играет роль в связывании поверхностного слоя слизи. Гликокаликс является не только диффузионным барьером, препятствующим проникновению бактерий, но и специфическим барьером, определяющим проницаемость пищевых веществ. Благодаря ему апикальная мембрана практически недоступна для высокомолекулярных соединений типа ксенобиотиков. В то нее время гликокаликс обеспечивает высокую проницаемость для молекул пищевых веществ, так как в его пространстве адсорбированы пищеварительные ферменты панкреатического происхождения.
От состояния гликокаликса и содержащихся в нем ферментов могут зависеть многие нарушения функций желудочно-кишечного тракта. В частности, нарушение адсорбции панкреатических ферментов имеет значение в формировании явлений малнутриции, а атрофия гликокаликса может явиться причиной повреждающего действия токсических агентов химуса на липопротеиновую мембрану.
Быстрое обновление гликокаликса обеспечивает эффективное функционирование щеточной каймы как пористого реактора, так как благодаря сбрасыванию «зрелого» гликокаликса создается своеобразный эффект постоянной очистки пор. Недавно показано активное сбрасывание гликокаликса в результате активации фибриллярных структур микроворсинок терминальной сети.
Следует подчеркнуть, что гликокаликс представляет собой универсальную структуру и встречается у клеток всех организмов от амебы до человека, включая эритроциты и нейроны. В то же время гликокаликс характеризуется вариабельностью свойств не только в клетках разного типа, но и в одной клетке. Например, он сильно развит на апикальной поверхности микроворсинок кишечных клеток, менее — на их латеральной поверхности и лишь биохимически (но не структурно) определяется на базальной мембране.
Актомиозиновый комплекс. Сократительные структуры характерны для возбудимых тканей (мышечной и нервной) и контролируют форму клеток. Однако оказалось, что эти структуры присутствуют в самых различных клетках. Недавно некоторые блоки сократительной системы описаны в невозбудимых клетках, и в том числе в кишечных.
На рис. 27 представлена сократительная система апикальной части кишечной клетки. Как можно видеть, сердцевина микроворсинки занята фибриллярными структурами, которые достигают апикальной области цитоплазмы, где сливаются с поперечной фибриллярной структурой — терминальной сетью. Последняя, согласно существующим представлениям, выполняет функцию опоры для щеточной каймы, принимает участие в регуляции подвижности мембраны, а также разделяет клетку на два компартмента — щеточнокаемный и цитозольный.
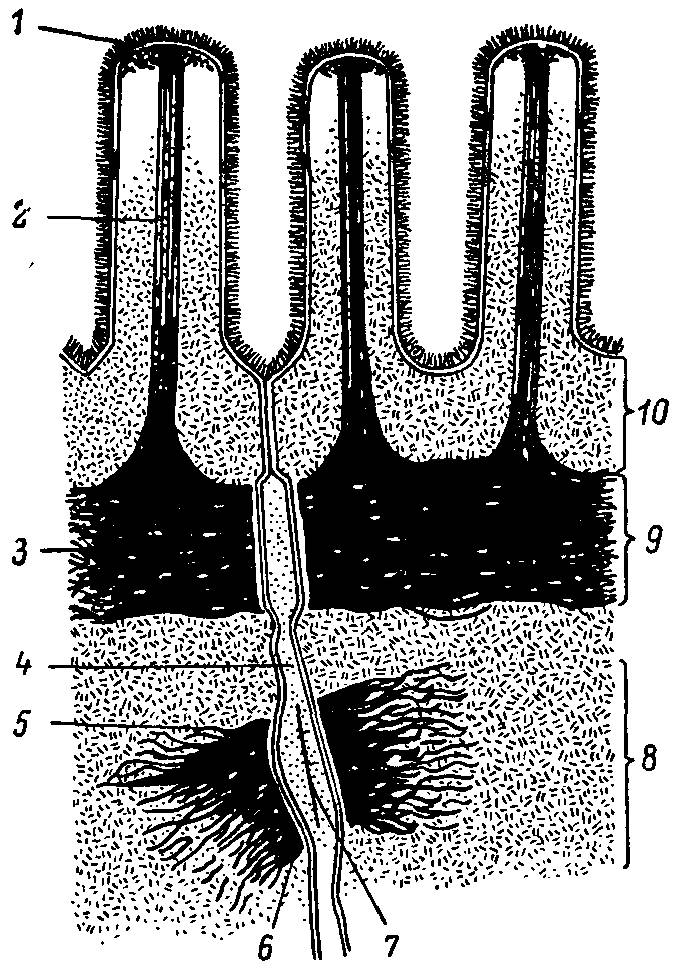
Рис. 27. Схема мембраны микроворсинок и апикальных соединительных комплексов латеральной мембраны кишечных клеток.
1 — гликокаликс; 2 — микрофиламенты; 3 — терминальная сеть; 4 — межклеточное пространство; 5 — тонофиламенты; в — плотное пятно; 7 — промежуточная плотная линия; 8 — десмосома; 9 — промежуточный контакт; 10 — плотный контакт.
Возникает вопрос о функциях сократительных структур в невозбудимых клетках. Нами еще в начале 60-х гг. было высказано предположение, получившее подтверждение, что благодаря актомиозиновому комплексу реализуются расслабления и сокращения микроворсинок, что может существенно влиять на скорость мембранного пищеварения и всасывания. Недавно выявлена еще одна функция сократительного аппарата кишечных клеток — контроль их десквамации, обеспечивающий гомеостатирование барьерных свойств эпителиального пласта.
Наличие сократительных белков в различных клетках немышечной ткани говорит об их универсальности.
При этом актины немышечных и мышечных клеток сходны по своей структуре. Вероятно, актомиозиновый комплекс, присутствующий во всех эукариотических клетках и участвующий в двигательных процессах, включен в функционирование как возбудимых, так и невозбудимых тканей. Актомиозиновые структуры обнаружены также у большинства растений, у высших и низших грибов. Наличие актомиозиновых белков, по-видимому, служит той эволюционной основой, благодаря которой периодически возникают уникальные способности двигательного аппарата. Примером этому могут служить насекомоядные растения. Возникновение у них двигательных функций не кажется теперь столь неожиданным, так как присутствие сократительных структур характерно для клеток всех типов.
В сократительном блоке принцип регуляции является общим и мало меняется на протяжении эволюции от примитивных грибов до человека. Во всех случаях сокращение связано с активирующим действием Ca2+ и АТФазной активностью миозина. Характерно варьирование компонентов актомиозинового комплекса, при котором один из них может выпадать. Как правило, в филогенезе наиболее стабилен актиновый компонент, тогда как миозиновый по своему составу варьирует в широких пределах, так же как и их соотношения.
Гидролитические ферменты апикальной мембраны кишечных клеток. Эти ферменты — один из основных типов функциональных блоков в системе, выполняющей пищеварительные и транспортные функции в организме (т.е. в тонкой кишке). Особое значение имеют сравнительные исследования клеток тонкой кишки и почки. Эти органы, выполняющие различные физиологические функции, обладают такими идентичными ферментами, как мальтаза, аминопептидазы, эндопептидаза, щелочная фосфатаза, трегалаза и др. (табл. 7). Недавно из щеточнокаемных мембран клеток почки и плаценты выделены такие одноименные ферменты, как аланинаминопептидаза, мальтаза, сахараза, щелочная фосфатаза и др. Многие из этих ферментов обнаружены в мембранах клеток печени и поджелудочной железы, в кровеносной системе и т.д. В некоторых случаях удается показать близость или даже идентичность таких ферментов. Так, свойства щелочной фосфатазы клеток печени, почки, тонкой кишки, плаценты и сыворотки крови обладают значительным сходством. Обнаружено также сходство в иммунохимических характеристиках аланинаминопептидазы клеток поджелудочной железы, почки и печени человека.
Ферменты мембраны микроворсинок клеток почки и тонкой кишки млекопитающих
Тривиальное наименование фермента Наименование по КФ и № Локализация Аминопептидаза Аминопептидаза (микросомная), 3.4.11.2 Присутствует в почке и кишке Аминопептидаза А Аспартатаминопептидаза, 3.4.11.7 То же Дипептидилпептпдаза IV Дипептидил пептидаза, 3.4.14.— или 3.4.21.— » Эндопептидаза Нейтральна я пептидаза, 3.4.24.11 » Глутамилтрансфераза Глутамилтрансфераза, 3.2.2.2. » Сахараза-изомальтаза Саха раза-a-D-гликогидролаза, олиго-1,6-глюкозидаза, 3.2.1.48, 3.2.1.10 Присутствует только в кишке Мальтаза (глюкоамилаза) a-D-Глюкозидаза, 3.2.1.20 Присутствует в кишке и почке некоторых видов Трегалаза а-a-Трегалаза, 3.2.1.28 Присутствует в почке и кишке Лактаза-гликозилкерамидаза ?-D-Галактозидаза, 3.2.1.23, 3.2.1.62 Присутствует только в кишке Щелочная фосфатаза Щелочная фосфатаза, 3.1.3 1 Присутствует в почке и кишкеОдни и те же ферменты, например щеточноrаемные гидролазы, могут присутствовать не только в мембране микроворсинок щеточной каймы, но и в латеральных и базальных мембранах, мембранах различных внутриклеточных органелл и даже в цитоплазме. Если присутствие щеточнокаемных ферментов в базолатеральной мембране свидетельствует об одном из физиологических путей от места синтеза к области их действия, то другая локализация, возможно, является полностью или частично результатом побочных эффектов. Однако
такие эффекты могут быть исходным материалом для формирования новых типов функций. Действительно, ферменты мембраны микроворсинок щеточной каймы, накапливаясь во внутриклеточных структурах, могут давать неожиданные физиологические эффекты.
Наконец, ранее предполагалось, что кишечные клетки отличаются от клеток других типов наличием мембранного пищеварения и отсутствием сократительных структур. Однако, как отмечено выше, пищеварительными свойствами обладают не только кишечные клетки, но и, в частности, клетки кровеносных сосудов. В начале 60-х гг. нами выявлено, что крахмал, проходя через сосуды пищеварительной системы и ряда других органов лягушки, подвергается значительному гидролизу. При этом расщепление крахмала обусловлено преимущественно ферментами, связанными со структурами периферического кровообращения. В 1980 г. нами совместно с коллегами из Военно-медицинской академии им. С. М. Кирова и Института физики АН СССР (Красноярск) показано, что мембранный гидролиз имеет место в ряде органов, в том числе в сосудистой системе кишечника, а также задних конечностей собак.
Таким образом, мембранное пищеварение и пищеварительные свойства мембран — не уникальное свойство кишечных клеток, а характерное для многих соматических клеток организма. Как продемонстрировано выше, типичные гидролазы мембраны микроворсинок кишечных клеток обнаружены в плазматических мембранах клеток многих непищеварительных органов, где эти ферменты, вероятно, выполняют другие функции.
Пищеварительные ферменты на всех этапах эволюции обладают примерно одинаковыми кинетическими константами и молекулярными характеристиками. Это справедливо, в частности, для таких универсальных пищеварительных ферментов, как кислые протеазы, амилазы и др., возникших на ранних стадиях эволюции и обнаруженных у наиболее примитивных форм, а также сохранившихся у высших организмов и растений. Даже когда ферменты распространены но столь широко, как, например, сериновые протеазы, их молекулярная структура и функции если не идентичны, то весьма близки. Что касается кислых протеаз, обнаруженных у микроорганизмов, высших растений, животных и человека, то, по-видимому, они возникли путем дупликации и слияния генов из общего предшественника.
Именно анализ эволюции пищеварительных ферментов, точнее, их стабильности, или консервативности в ходе эволюции наряду с очевидной адаптивностью экзотрофических процессов, явился одним из краеугольных камней для представлений о рекомбинации универсальных функциональных блоков как важном механизме эволюции.
Таким образом, мембранные пищеварительные ферменты распространены чрезвычайно широко, причем между мембранными ферментами различных органов существует большое сходство.
Транспортеры и ферментно-транспортные комплексы кишечных клеток. Многие общие и специализированные функции клеток различных типов связаны с деятельностью транспортных систем, обеспечивающих активную и пассивную, селективную и неселективную проницаемость. Предполагается, что все многообразие разновидностей молекулярного специфического транспорта через мембрану, включая активный транспорт против градиента концентраций, реализуется с помощью нескольких типов молекулярных машин — насосов, переносчиков, каналов и пор. Одним из примеров универсальных функциональных блоков мембраны кишечных клеток служат транспортеры, реализующие перенос нутриентов через мембрану. Так, перенос глюкозы осуществляется в результате взаимодействия этого вещества с олигомерным белком, представляющим собой скорее всего трансмембранную олигомерную структуру с молекулярной массой около 100 000 даль-тон. Эта структура путем изменения либо конформации, либо своего положения способна переносить глюкозу через гидрофобный фосфолипидный матрикс мембраны в цитоплазму. В клетках кишечника, мозга, мышц, почек, жировой ткани и других транспорт глюкозы, как и аминокислот, зависит от натрия. Более того, Na+-зaвисимым является трансмембранный перенос этого моносахарида у всех организмов — от дрожжей до высших эукариотов, включая простейших, растения, животных и человека. Другой широко распространенный блок, деятельность которого не зависит от натрия, — комплекс, состоящий из ферментной и транспортной частей. Один из компонентов комплекса реализует гидролиз олигомера, например олигосахарида или олигопептида, до мономеров, например глюкозы или аминокислот, а другой — Na+-независимый перенос продуктов реакции с ферментной части блока на транспортную без выхода в водную фазу. Такой пищеварительно-транспортный блок универсален и обнаружен у всех организмов, находящихся на различных ступенях эволюции. У высших животных и человека он описан не только в мембранах клеток пищеварительного аппарата, но и в соматических клетках.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Бактериальная клетка в цифрах
Бактериальная клетка в цифрах Благодаря биофизике — одной из отраслей науки, с которой мы уже познакомились в начале этой главы, — были получены весьма интересные данные. Возьмем, например, шаровидную бактериальную клетку диаметром 0,5 мкм. Поверхность такой клетки
ГРУДНАЯ КЛЕТКА
ГРУДНАЯ КЛЕТКА Форма грудной клетки изменяется в зависимости от конституционального типа собаки, степени ее развития и возраста. Грудная клетка, вмещающая дыхательные органы, сердце и главнейшие кровеносные сосуды, должна быть объемистой. Объем груди обусловлен длиной,
Глава 4. Сложная клетка
Глава 4. Сложная клетка Ботаник — это тот, кто умеет давать одинаковые названия одинаковым растениям и разные названия разным, причем так, чтобы в этом мог разобраться каждый”, - писал великий шведский систематик Карл Линней (сам ботаник). Это определение может поразить
Клетка-ловушка
Клетка-ловушка Тебе понадобятся: клетка-ловушка, приманка (зерна, сыр, хлеб, колбаса), доска или черепицаДлительность опыта: 1–2 дня.Время проведения: поздняя осень — ранняя весна. Твои действия:• Купи клетку-ловушку любого типа или сделай ее сам. Для этого возьми
Кишечная непроходимость
Кишечная непроходимость Кишечная непроходимость возникает в результате запоров, из-за глистов или проглоченных посторонних предметов, а также вследствие заворота кишок.При кишечной непроходимости у собаки появляется рвота, пропадает аппетит. Животное находится в
5.6. Концепция универсальных функциональных блоков и кишечная гормональная система
5.6. Концепция универсальных функциональных блоков и кишечная гормональная система Недавно, когда среди гормонов, продуцируемых эндокринными клетками тонкой кишки, оказались гормоны, характерные для гипоталамуса, гипофиза, щитовидной железы, коры надпочечников и т.д.,
Глава 2. Клетка
Глава 2. Клетка ТЕМЫ• История изучения клетки. Клеточная теория• Химический состав клетки• Строение эукариотической и прокариотической клеток• Реализация наследственной информации в клетке• ВирусыУдивительный и загадочный мир окружает нас, жителей планеты,
12. Прокариотическая клетка
12. Прокариотическая клетка Вспомните!В чём заключаются принципиальные отличия в строении прокариотических и эукариотических клеток?Какова роль бактерий в природе?Разнообразие прокариот. Царство прокариот в основном представлено бактериями, наиболее древними
Глава 6. Кишечная гормональная система и трофика организма
Глава 6. Кишечная гормональная система и трофика организма 6.1. Вводные замечания Одно из наиболее существенных различий теорий сбалансированного и адекватного питания заключается в том, что в соответствии с последней ассимиляция определяется не только поступлением
7.3. Специфическое динамическое действие пищи и кишечная гормональная система
7.3. Специфическое динамическое действие пищи и кишечная гормональная система Вся совокупность данных позволяет считать классические представления о СДД пищи слишком упрощенной интерпретацией сложного и многообразного процесса, точнее совокупности процессов,
8.4. Аппетит и кишечная гормональная система
8.4. Аппетит и кишечная гормональная система В течение последних десятилетий получено много данных о важной роли кишечных гормонов в регуляции потребления пищи. Кроме того, в течение последней четверти века представления о регуляции аппетита изменились в связи с
Глава 6. Кишечная гормональная система и трофика организма
Глава 6. Кишечная гормональная система и трофика организма 6.1. Вводные замечания Одно из наиболее существенных различий теорий сбалансированного и адекватного питания заключается в том, что в соответствии с последней ассимиляция определяется не только поступлением
7.3. Специфическое динамическое действие пищи и кишечная гормональная система
7.3. Специфическое динамическое действие пищи и кишечная гормональная система Вся совокупность данных позволяет считать классические представления о СДД пищи слишком упрощенной интерпретацией сложного и многообразного процесса, точнее совокупности процессов,
8.4. Аппетит и кишечная гормональная система
8.4. Аппетит и кишечная гормональная система В течение последних десятилетий получено много данных о важной роли кишечных гормонов в регуляции потребления пищи. Кроме того, в течение последней четверти века представления о регуляции аппетита изменились в связи с