8.4. Аппетит и кишечная гормональная система
8.4. Аппетит и кишечная гормональная система
В течение последних десятилетий получено много данных о важной роли кишечных гормонов в регуляции потребления пищи. Кроме того, в течение последней четверти века представления о регуляции аппетита изменились в связи с открытием специализированных гормональных влияний на потребление пищи.
8.4.1. Арэнтерин
В 1960 г. нами (Уголев, 1960б) было продемонстрировано, что экстракты двенадцатиперстной кишки, но не желудка, печени, мышц и селезенки, вызывают специфическое торможение аппетита (рис. 8.1). После введения экстракта двенадцатиперстной кишки поведение голодных крыс напоминало поведение сытых животных. Снижение потребления пищи наблюдалось после введения таких экстрактов как сытым, так и голодным животным. На основании этих данных можно допустить, что существует кишечный гормон или фактор, осуществляющий торможение аппетита, т. е. торможение той важнейшей реакции, которая характеризует изменение поведения человека и животных после удовлетворения пищевых потребностей. В 1962 г. (Уголев, 1962) были получены сведения, которые позволили предположить, что такой фактор, скорее всего, является пептидом. Наличие дуоденального фактора, вызывающего сытость, который был назван нами арэнтерином (аппетитрегулирующим энтерином, т. е. кишечным гормоном), так же как и его пептидная природа получили подтверждение во многих работах. Все же следует отметить, что до сих пор арэнтерин в чистом виде не получен.
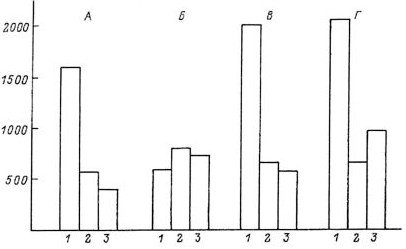
Рис. 8.1. Потребление пищи предварительно голодавшими крысами после введения раствора Рингера (А), экстрактов двенадцатиперстной кишки (Б), желудка (В) и селезенки (Г) (по: Уголев, 1978).
По горизонтали — время от начала кормления: 1 — 0–30 мин, 2 — 30–60 мин, 3 — 60–120 мин; по вертикали — количество потребляемой пищи (мг).
Попытки охарактеризовать арэнтерин были предприняты группой чл. — корр. АН СССР А. С. Хохлова в Институте биоорганической химии им. М. М. Шемякина АН СССР и в нашей лаборатории. В середине 70-х годов обеими группами было продемонстрировано, что арэнтерин — это белок с молекулярной массой около 100000 (обзоры: Уголев, 1978, 1985).
8.4.2. Другие кишечные гормоны
В 70-х годах было обнаружено, что многие известные гормоны желудочно-кишечного тракта, в частности гастрин и секретин, не оказывают влияния на аппетит. В отличие от них холецистокинин вызывает сильное торможение пищевой активности. Первым, по-видимому, продемонстрировал специфическое влияние холецистокинина на потребление пищи X. Купманс с сотрудниками (Koopmans et al., 1972). В цикле исследований группы Дж. Смита (Smith et al., 1974) на крысах было показано, что холецистокинин может вызывать торможение аппетита. Авторы, однако, оценивали этот эффект, скорее, как фармакологический, чем физиологический. у обезьян также обнаружено торможение потребления пищи после введения холецистокинина. У человека потребление пищи зависит от дозы введенного холецистокинина и от способа его введения. Так, внутривенное введение гормона в дозе 0.5 ед/кг приводит к торможению аппетита, а постепенное внутривенное введение в дозе 1 ед/кг, напротив, вызывает его стимуляцию (Sturdevant, Goetze, 1976). Прямое действие холецистокинина на центры питания и другие нервные центры показано в электрофизиологическом исследовании Н. Дафни и Е. Джекобсона (Dafny, Jacobson, 1975). Наконец, продемонстрировано, что после введения глюкагона возрастает электрическая активность центра сытости. (Более подробно эти вопросы освещены в монографии: Уголев, 1978.).
В 80-е годы интенсивная работа в этом направлении продолжалась. На человеке и животных было вновь продемонстрировано, что в передаче сигнала насыщения большую роль играют кишечные гормоны. Суммируя полученные за эти годы данные, можно сказать, что экзогенный холецистокинин, в том числе его синтетический октапептид холецистокинин-8, вызывает снижение потребления пищи, которое зависит от дозы гормона и места его введения (периферического или центрального). При этом для развития эффекта насыщения необходим сульфатированный тирозин в 7-й позиции С-терминального конца цепи октапептида холецистокинина. Показана также физиологическая специфичность холецистокинина: он ингибирует потребление пищи, но не воды (Smith, 1983; Gibbs, Smith, 1986, и др.).
В последние годы дискутируется вопрос о периферическом и центральном механизмах действия холецистокинина на аппетит. (Физиологический и фармакологический эффекты гормона дифференцировать достаточно трудно.)
Так как систематические инъекции холецистокинина приводят к резкому изменению пищевого поведения, то предполагается, что гормон должен прямо или опосредованно вызывать изменения функционального состояния мозговых структур. Считается, что основным путем передачи информации после введения экзогенного холецистокинина служат афферентные волокна абдоминальной вагусной системы, с рецепторами которой связывается гормон. Кроме того, гормон может связываться и с рецепторами пилорического сфинктера желудка.
Для объяснения эффекта эндогенного холецистокинина привлекается гипотеза, согласно которой освобождение этого гормона, вызывая чувство насыщения, определяет окончание акта еды, следствием чего является соответствующее поведение. По-видимому, эндогенный холецистокинин может прямо влиять на функции мозга, вызывая торможение аппетита. Принято, что основную роль в этом эффекте играют вентромедиальная и латеральная области гипоталамуса.
В последние годы продемонстрировано, что такие кишечные пептиды, как глюкагон, соматостатин и бомбезин, также вызывают торможение потребления пищи, но в меньшей мере, чем холецистокинин. В отличие от этих гормонов секретин, гастрин и ГИП не обладают таким эффектом.
Подробно история развития проблемы и ее современное состояние освещены з ряде детальных сводок (Smith, 1982, 1983, 1984; Gibbs, Smith, 1986, 1988; Murphy et al., 1988; Кассиль, 1990, и др.).
Наконец, к кишечным пептидным гормонам, ингибирующим потребление пищи, относится также сатиетин (обзор: Замбржицкий, 1989).
Следует отметить, что влияние на аппетит холецистокинина, глюкагона, энтерогастрона и других гормонов, по-видимому, является важной, но скорее дополнительной (а не основной) функцией. По крайней мере в некоторых случаях можно думать об их фармакологическом, а не физиологическом действии. Эти же факты давали возможность предполагать, что среди кишечных гормонов, возможно, существуют такие, которые обладают преимущественно нейротропным действием, и их аппетиттормозящие эффекты будут выражены более отчетливо, чем у известных гормонов. Действительно, нами было продемонстрировано, что арэнтерин влияет на потребление пищевых веществ более эффективно, чем холецистокинин и секретин. В частности, при внутрибрюшинном введении препарата арэнтерина в дозе 0.1 мг на 100 г массы тела крысы наблюдается резкое торможение потребления раствора глюкозы (рис. 8.2). После введения заведомо большей дозы секретина (0.5 мг на 100 г массы тела) торможения аппетита не происходит. Введение холецистокинина также в дозе 0.5 мг на 100 г массы тела вызывает сравнительно слабый и кратковременный эффект (рис. 8.3) (Уголев, 1978).
Итак, эндокринные клетки тонкой кишки, по-видимому, продуцируют более чем один фактор, способный понижать пищевую возбудимость. В одних случаях некоторые из этих факторов — хорошо известные гормоны, проявляющие новые стороны физиологической (или фармакологической) активности, в других случаях — это «новые» гормоны. К числу последних относится арэнтерин. Заслуживает внимания, что арэнтерин обладает большей продолжительностью действия, чем холецистокинин. В этом смысле арэнтерин вообще отличается от большинства кишечных гормонов, эффект которых непродолжителен. Это означает, что кишечная гормональная система способна не только к кратковременным, но и более продолжительным регуляторным эффектам.
Рассмотрим более обобщенно влияние желудочно-кишечных гормонов на потребление пищи. Следует различать два типа эффектов: 1) прямые (гипоталамотропные) эффекты, в том числе арэнтерина и холецистокинина, на структуры гипоталамуса; 2) косвенные эффекты, которые, по-видимому, следует учитывать в комплексе реакций, связанных с нормальным насыщением. В частности, согласно дегидратационной теории регуляции аппетита, физиологическая дегидратация тканей, вызванная секрецией главных пищеварительных органов, служит одной из причин понижения аппетита и возникновения физиологической жажды. С этой точки зрения, можно думать, что в регуляции аппетита опосредованно участвуют: секретин, который стимулирует секрецию жидкой части поджелудочного сока, и гастрин, который вызывает значительную по объему секрецию желудочного сока. Кроме того, растяжение желудка, ацидификация желудка и верхних отделов двенадцатиперстной кишки являются также источником нервных сигналов, передаваемых главным образом по блуждающим нервам.
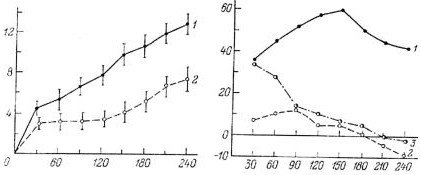
Рис. 8.2. Потребление глюкозы после внутрибрюшинного введения раствора Рингера и препарата арэнтерина (по: Уголев, 1978).
По оси абсцисс — время после потребления глюкозы (мин); по оси ординат — количество выпитого 40 %-ного раствора глюкозы (мл). 1 — после введения раствора Рингера, 2 — после введения арэнтерина.
Рис. 8.3. Торможение потребления глюкозы после внутрибрюшинного введения препаратов арэнтерина, секретина и холецистокинина (по: Уголев, 1978).
По оси абсцисс — время после потребления глюкозы (мин); по оси ординат — торможение потребления 4-%-ного раствора глюкозы (% по отношению к исходному уровню). 1 — после введения арэнтерина; 2 — после введения секретина; 3 — после введения холецистокинина.
Особого рассмотрения заслуживает роль кишечной гормональной системы в потреблении пищи в связи с термостатической теорией регуляции аппетита. Ряд данных позволяет считать, что СДД пищи и изменение внутренней, в частности гипоталамической, температуры могут быть важным компонентом нормального насыщения. Так, показано, что повреждения ростральной области гипоталамуса, связанной с регуляцией как температуры, так и потребления пищи, приводят к нарушению обеих этих функций (обзор: Hamilton, 1967). Здесь необходимо напомнить о существовании обнаруженных нами компонентов СДД пищи (см. гл. 7), контролируемых кишечной гормональной системой и действием желудочного химуса при его прохождении через двенадцатиперстную кишку. Понижение пищевого возбуждения, связанное с интенсификацией обмена, и повышение внутренней температуры также в определенной мере обусловлены включением кишечного гормонального комплекса. В целом прохождение пищи через верхние отделы пищеварительного аппарата вызывает выделение ряда гормонов желудочно-кишечного тракта, прямо или косвенно влияющих на переход от голодного состояния к сытому и во многих случаях к появлению питьевых реакций. К гормонам, вызывающим физиологическую дегидратацию, которая благоприятствует понижению пищевого возбуждения и повышению питьевого, относятся гастрин и секретин. Дегидратация может влиять на пищевой центр как через периферические, так и через центральные осморецепторы.
В свете обсуждаемых данных обращает на себя внимание недавнее исследование, на основании которого высказано предположение, что мотилин играет важную роль в контроле моторной деятельности не только у собак и людей, но и у свиней (Bull, Sissons, 1988).
Совершенная регуляция потребления пищи включает в себя два типа регуляторов, различающихся по своим временным характеристикам: быстродействующие (кратковременные) и длительно действующие (долговременные). Как правило, регуляция первого типа обеспечивает срочные, но не вполне точные реакции, тогда как медленные регуляции обеспечивают точное соответствие между потребностью в калориях и их поглощением.
Таким образом, если существует несколько типов кишечных гормонов, влияющих на аппетит (а их должно быть несколько, в том числе арэнтерин и холецистокинин), то с их помощью реализуются как более быстрые, так и более медленные типы регуляции аппетита, или потребления пищи (рис. 8.4).
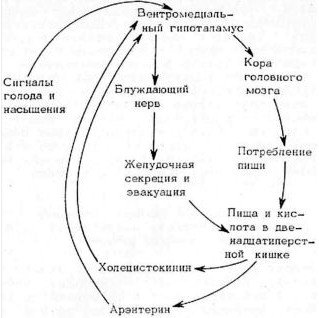
Рис. 8.4. Схема нейрогуморальной регуляции аппетита с участием арэнтерина (по: Уголев, 1978).
До сих пор мы рассматривали влияние кишечной гормональной системы на пищевой центр, сузив эту проблему до регуляции потребления пищевых веществ. В действительности речь идет о том, что кишечная гормональная система путем изменения мотиваций участвует в переключении целенаправленного поведения, характерного для голодного животного, на поведение, свойственное сытому животному. Поэтому можно думать, что чем дальше будет развиваться эта проблема, тем больше места в ней будет занимать область нейротропных и психотропных влияний кишечной гормональной системы.
Как отмечено выше, Н. Дафни Е. Джекобсон (Dafny, Jacobson, 1975) показали, что гормоны желудочно-кишечного тракта, продуцируемые при потреблении пищи, индуцируют насыщение посредством изменения электрической активности центральной нервной системы. По мнению авторов, центр аппетита взаимодействует с вышележащими центрами, которые могут влиять на тонус сосудов, моторику пищеварительного аппарата и секреторную активность. Предполагается, что холецистокинин может действовать как модулятор одного из известных химических мессенджеров. Вместе с тем эти данные подтверждают высказанную нами мысль, что эффекты гормонов желудочно-кишечного тракта должны охватывать в конце концов более значительную сферу реакций, чем только контроль над пищеварительными процессами.
В целом приведенные сведения свидетельствуют, что кишечная гормональная система может играть важную роль не только в контроле обмена веществ за пределами пищеварительного аппарата, но и в управлении поведенческими реакциями, прямо или косвенно связанными с регуляцией потребления пищи.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
7. ВКУСОВАЯ СИСТЕМА
7. ВКУСОВАЯ СИСТЕМА В процессе эволюции вкус формировался как механизм выбора или отвергания пищи. Выбор предпочитаемой пищи отчасти основан на врождённых механизмах, но в значительной мере зависит от связей, выработанных в онтогенезе. Вкус, так же как и обоняние, основан
2.1. Что такое система?
2.1. Что такое система? Термин «система» обычно применяется для того, чтобы указать на собранность, организованность группы элементов и отграниченность её от других групп и элементов. Давалось множество определений системы, которые характеризовали её, выделяли из
Возрастные изменения и гормональная регуляция
Возрастные изменения и гормональная регуляция Два обстоятельства в исследовании возрастных изменений обращают на себя внимание: 1) некоторая общность проявлений отдаленной радиационной и возрастной патологии; 2) взаимосвязь многих проявлений возрастной патологии с
11.2. АППЕТИТ
11.2. АППЕТИТ Под понятием «аппетит» разные авторы понимают не вполне совпадающие, хотя и имеющие много общего, феномены пищевого поведения. Для одних аппетит сродни голоду, и они измеряют его количеством добровольно поглощаемой пищи. Для других аппетит — это стремление
Волчий аппетит
Волчий аппетит Rhone-Poulenc, филиалом которой является компания Pasteur-Merileux — производитель сывороток и вакцин, опубликовала в прессе о своем решении приобрести 100 % пакет акций своего американского филиала Rhone-Poulenc Rorer.Эта транснациональная компания французского
Система FCI:
Система FCI: В соответствии с решением Генерального собрания FCI в Иерусалиме в 1987 году принято следующее деление зарегистрированных пород собак на группы, которое вступило в силу с 1 января 1990 года.1 группа — овчарки (пастушьи и скотогонные собаки, за исключением
У кого какой аппетит
У кого какой аппетит Аппетит у насекомых самый разный. И количество поедаемой пищи часто не связано ни с размерами, ни с активностью, хотя она, безусловно, имеет немалое значение. Те, кто поедает пищу малопитательную, уничтожает ее подчас в огромных количествах. Очень
Кишечная непроходимость
Кишечная непроходимость Кишечная непроходимость возникает в результате запоров, из-за глистов или проглоченных посторонних предметов, а также вследствие заворота кишок.При кишечной непроходимости у собаки появляется рвота, пропадает аппетит. Животное находится в
5.2. Кишечная клетка
5.2. Кишечная клетка Схема кишечной клетки представлена на рис. 26. Известно, что численность кишечных клеток составляет 1010, а соматических клеток взрослого человека— 10 15. Следовательно, одна кишечная клетка обеспечивает питание около 100 000 других клеток. Такая
5.6. Концепция универсальных функциональных блоков и кишечная гормональная система
5.6. Концепция универсальных функциональных блоков и кишечная гормональная система Недавно, когда среди гормонов, продуцируемых эндокринными клетками тонкой кишки, оказались гормоны, характерные для гипоталамуса, гипофиза, щитовидной железы, коры надпочечников и т.д.,
9. Нервная система
9. Нервная система Общие понятия. Нервная система является очень сложной и своеобразной по своему строению и функциям системой организма. Ее назначение — устанавливать и регулировать взаимоотношение органов и систем в организме, связывать все функции организма в
Глава 6. Кишечная гормональная система и трофика организма
Глава 6. Кишечная гормональная система и трофика организма 6.1. Вводные замечания Одно из наиболее существенных различий теорий сбалансированного и адекватного питания заключается в том, что в соответствии с последней ассимиляция определяется не только поступлением
7.3. Специфическое динамическое действие пищи и кишечная гормональная система
7.3. Специфическое динамическое действие пищи и кишечная гормональная система Вся совокупность данных позволяет считать классические представления о СДД пищи слишком упрощенной интерпретацией сложного и многообразного процесса, точнее совокупности процессов,
8.4. Аппетит и кишечная гормональная система
8.4. Аппетит и кишечная гормональная система В течение последних десятилетий получено много данных о важной роли кишечных гормонов в регуляции потребления пищи. Кроме того, в течение последней четверти века представления о регуляции аппетита изменились в связи с
Глава 6. Кишечная гормональная система и трофика организма
Глава 6. Кишечная гормональная система и трофика организма 6.1. Вводные замечания Одно из наиболее существенных различий теорий сбалансированного и адекватного питания заключается в том, что в соответствии с последней ассимиляция определяется не только поступлением
7.3. Специфическое динамическое действие пищи и кишечная гормональная система
7.3. Специфическое динамическое действие пищи и кишечная гормональная система Вся совокупность данных позволяет считать классические представления о СДД пищи слишком упрощенной интерпретацией сложного и многообразного процесса, точнее совокупности процессов,