9.6. Трофические цепи и экология
9.6. Трофические цепи и экология
Одним из следствий развиваемого нами трофологического подхода (см. гл. 1) является признание того, что процветание вида во многом определяется его положением в трофической цепи. Это положение обеспечивается эффективностью взаимодействий не только с предшествующими, но и с последующими членами трофической цепи. Другими словами, существенную роль играет не только источник питания и его эффективное поглощение, но и поедаемость данного члена экосистемы.
Трофическая цепь возможна лишь при соответствии структур предшествующего трофического звена, т. е. трофического субстрата, ферментам следующего трофического звена, утилизирующего предыдущее как пищевое вещество. Такое соответствие должно быть и между ферментами данного звена трофической цепи и его же собственными субстратами, что необходимо для реализации различных функций внутри данного организма.
Во взаимоотношениях хищник-жертва, независимо от того, рассматриваются ли два вида животных, или растительноядное животное и поедаемое растение, жертва как вид может сохраниться лишь при наличии у нее укрытия и средств активной и пассивной защиты. Нередко два последних эффекта достигаются с помощью ядов.
Животные для защиты и нападения часто используют яды и токсины. В качестве таких примеров можно привести змей, пауков, скорпионов, пчел, ос и др. (см. обзоры: Rice, 1978; Сравнительная физиология…, 1978; Schmidt-Nielsen, 1982; Odum, 1986, и др.). У многих организмов яд служит не только средством защиты от врагов, но и средством поражения добычи. Интересные средства защиты применяются некоторыми многоножками, которые продуцируют синильную кислоту, или жуками-бомбардирами, выбрасывающими в виде защиты струю аэрозоля с температурой 10 °C. Широко распространены растительные яды, предохраняющие растения (первичные продуценты трофической цепи) от поедания. Однако существуют примеры химических взаимодействий, при которых хищник может использовать жертву без отрицательных для себя последствий. Так, гусеницы бабочки-капустницы могут питаться капустой и горчицей без вредных для себя последствий, тогда как эти же крестоцветные ядовиты для гусениц других бабочек. Более того, горчичные масла этих растений представляют собой аттрактанты, привлекающие бабочек для откладки яиц. Один из способов защиты некоторых животных заключается в аккумуляции токсических веществ растений, что делает этих животных несъедобными для врагов. Таким примером может служить американская бабочка-данаида, которая, потребляя в пищу обычный ваточник, синтезирующий гликозиды, становится вредной для своих хищников.
В ряде случаев растения продуцируют вещества, действующие на эндокринную систему насекомых и позвоночных. Так, некоторые американские вечнозеленые растения вырабатывают сходные с ювенильным гормоном биологически активные вещества, которые тормозят размножение насекомых, останавливая переход последних во взрослое состояние. Некоторые пастбищные растения, в частности бобовые, синтезируют биологически активные вещества, обладающие эстрогенными эффектами. Эти вещества влияют на половые функции животных, нарушая цикл воспроизводства крупного рогатого скота и овец. Кроме того, фитоэстрогены могут регулировать численность диких полевых грызунов.
В некоторых случаях показана прямая зависимость между размножением животных и продуктивностью растений. Так, в Калифорнии при засухе ряд однолетних растений вырабатывает фитоэстрогены, ингибирующие появление потомства у перепелов. Напротив, при благоприятных условиях концентрация этих эстрогенов в растениях становится низкой, что способствует размножению птиц. Упомянутый ранее ваточник синтезирует сердечные гликозиды, которые оказывают действие на сердце и мозговые центры поедающих его животных, провоцируя рвоту.
Приспособление к питанию растениями, содержащими сердечные гликозиды, выработалось у некоторых насекомых в результате мутации, вызвавшей потерю чувствительности Na+,К+ — АТФазы к уабаину. В качестве примера можно вновь привести бабочку-данаиду, у которой этот фермент в отличие от АТФаз других организмов не чувствителен к уабаину. Эта точковая мутация — несомненно полезный селективный признак, так как данаида питается растениями, содержащими сердечные гликозиды.
Этих примеров вполне достаточно для иллюстрации сложности трофических отношений в природе между различными организмами и широким распространением ядовитых веществ.
Одним из примеров интересных путей эволюции могут быть насекомые с их изощренными механизмами нападения и умерщвления жертвы, ее обездвиживания и сохранения в живом, но неподвижном состоянии на протяжении длительных интервалов времени. Однако хотелось бы обратить внимание на другое обстоятельство. Во всех случаях убивающие и парализующие яды оказались агентами, действующими на определенные универсальные функциональные блоки (см. гл. 1), причем сами яды являются соединениями, достаточно широко распространенными в природе. Общность функциональных блоков настолько велика, что, хотя ветви насекомых и млекопитающих разошлись задолго до формирования систем свертывания и противосвертывания крови, у многих насекомых синтезируются биологически активные факторы, идентичные таковым млекопитающих. Примером может служить гепарин. Если насекомое питается тканевой жидкостью, то вводится гиалуронидаза, повышающая проницаемость тканей. Этот же фактор входит в физиологическую цепь регуляции вазопрессином некоторых функций почечных канальцев.
Наконец, существует еще один этап усвоения пищи в естественных условиях — индуцированный аутолиз (см. гл. 1).. Так, пауки для извлечения питательных веществ из жертвы прокалывают ее челюстями и вводят пищеварительные соки, которые, как предполага лось, растворяют ткани. Затеи пауки высасывают полученный раствор. Анализ в этих соках состава ферментов, обеспечивающих внеклеточное пищеварение, заставляет думать, что интерпретация данного феномена неправильна, так как внеклеточные ферменты реализуют преимущественно начальные этапы гидролиза. По всей вероятности, пауки наряду с пищеварительными ферментами вводят факторы, индуцирующие аутолиз. В этом случае образуется гидролизат, близкий к набору конечных продуктов расщепления, который и используется хищниками. Аналогичный механизм применяется и другими беспозвоночными, например жуками-плавунцами.
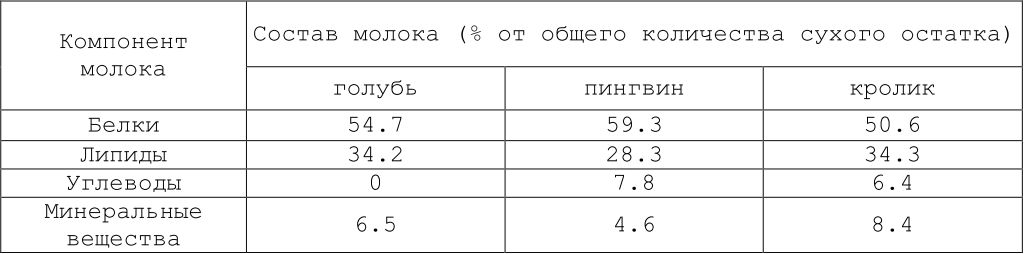
Рассмотрим еще один конкретный случай, за которым, однако, скрываются общие закономерности. У многих высших организмов ранние этапы онтогенеза обеспечиваются тем, что происходит прямой обмен макромолекулами и макромолекулярными функциональными комплексами между материнским и детским организмами. Такой обмен происходит при молочном питании у млекопитающих и имеет место при псевдомолочном питании у некоторых птиц. У голубя, и особенно у пингвина, вырабатывается питательная жидкость — так называемое молоко. Местом его образования у голубя служит зоб, и выработка молока стимулируется пролактином. У пингвина такое молоко образуется в пищеводе. Обращает на себя внимание близость состава птичьего молока и молока млекопитающих (табл. 9.2) (Prevost, Vilter, 1962). Ясно, что в обоих случаях молочное питание позволяет избежать случайностей, связанных с внешними обстоятельствами, и обеспечивает химическое гомеостатирование внутренней среды потомства в раннем постнатальном периоде.
Таблица 9.2. Состав «молока» голубя, императорского пингвина и молока кролика (по: Prevost, Vilter, 1962)
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Структурная организация цепи тканевого дыхания
Структурная организация цепи тканевого дыхания Компоненты дыхательной цепи во внутренней мембране михохондрий формируют комплексы:1. I комплекс (НАДН-КоQН2-редуктаза) – принимает электороны от митохондриального НАДН и транспортирует их на КоQ. Протоны транспортируются
7.3.6. Трофические связи
7.3.6. Трофические связи Трофические связи возникают, когда один вид питается другим – либо живыми особями, либо их мертвыми остатками, либо продуктами жизнедеятельности. И стрекозы, ловящие на лету других насекомых, и жуки-навозники, питающиеся пометом крупных копытных, и
ИСТОРИЧЕСКАЯ ЭКОЛОГИЯ
ИСТОРИЧЕСКАЯ ЭКОЛОГИЯ Невозможно понять экологию современности без знания экологии прошлого. Историческая экология — это «история растительности и ландшафтов» (Рэкхэм, 1998). За тысячи лет человек изменил многие среды обитания; в настоящее время многие ландшафты имеют
ЛАНДШАФТНАЯ ЭКОЛОГИЯ
ЛАНДШАФТНАЯ ЭКОЛОГИЯ Ландшафтная экология — новая дисциплина, которая изучает различные экологические процессы, происходящие на территориях, измеряемых гектарами и квадратными километрами.Такие огромные территории обычно состоят из отдельных фрагментов, как-то:
МОЛЕКУЛЯРНАЯ ЭКОЛОГИЯ
МОЛЕКУЛЯРНАЯ ЭКОЛОГИЯ В прессе часто появляются заметки о том, что торговцы дикими животными пытаются продавать запрещенные виды или продукты, изготовленные из вымирающих видов животных, прикрываясь торговлей вполне легальным товаром.Один из способов разрешения
ПОВЕДЕНЧЕСКАЯ ЭКОЛОГИЯ
ПОВЕДЕНЧЕСКАЯ ЭКОЛОГИЯ Почему некоторые животные проявляют альтруизм? Что мы имеем в виду под словом «поведение»?Основные положения поведенческой экологии заключаются в том, что существует генетический компонент поведения, который может регулироваться естественным
ХИМИЧЕСКАЯ ЭКОЛОГИЯ
ХИМИЧЕСКАЯ ЭКОЛОГИЯ Потеющие и плохо пахнущие ноги — это вовсе не смертельно и даже не опасно для здоровья, но только если вы не живете в странах, где распространена малярия. Комаров — переносчиков малярии привлекает запах различных химических веществ, которые выделяет
ЭКОЛОГИЯ
ЭКОЛОГИЯ В каком-то смысле наука экология такая же древняя, как человек. Люди всегда зависели от окружающего мира, от потребляемых ими животных и растений. Им нужно было знать, на каких животных охотиться, какие растения собирать и выращивать. Но самостоятельной научной
ЭКОЛОГИЯ МИКРООРГАНИЗМОВ
ЭКОЛОГИЯ МИКРООРГАНИЗМОВ Людей впечатляют большие размеры. Наверное поэтому, вспоминая о юрском периоде, мы в первую очередь представляем себе гигантских динозавров, когда-то «правивших» нашей планетой. Однако, если какие-то организмы и «управляют» Землей, то это
ЭКСПЕРИМЕНТАЛЬНАЯ ЭКОЛОГИЯ
ЭКСПЕРИМЕНТАЛЬНАЯ ЭКОЛОГИЯ Эксперименты играют решающую роль в науке. Они необходимы для проверки гипотез; при проведении экспериментов определенные факторы, интересующие ученых, подвергают различным изменениям, оставляя неизменными все другие факторы (или по крайней
9.6. Трофические цепи и экология
9.6. Трофические цепи и экология Одним из следствий развиваемого нами трофологического подхода (см. гл. 1) является признание того, что процветание вида во многом определяется его положением в трофической цепи. Это положение обеспечивается эффективностью взаимодействий
Звенья одной цепи
Звенья одной цепи В. П. Данилин решил узнать, как люди оценивают интервалы времени, прошедшие во сне. Он исходил из предположения, что адекватная оценка времени означает во всех случаях, что у человека в памяти фиксируется непрерывная последовательность событий,
Лев на золотой цепи
Лев на золотой цепи Даже царь зверей жил пленником у царей человеческих. В Египте сохранились древние тексты и барельефы, а на них фараоны с ручными львами. На одной из стен дворца в Карнаке изображен Рамзее II (годы его царствования 1324–1258 до нашей эры). Он во главе войска на