1.5. Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера
1.5. Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера
С деятельностью живых систем связана та часть поверхности Земли, которая объединена под названием биосферы. Биосфера, являющаяся самой крупной экосистемой, представляет собой открытую систему, т. е. получает и отдает энергию. Поток энергии направлен в одну сторону, причем часть поступающей солнечной энергии трансформируется в органическое вещество, а большая часть деградирует. Энергия может накапливаться, а затем снова освобождаться и экспортироваться. В отличие от энергии пищевые вещества, в том числе витальные биогенные элементы (углерод, азот, фосфор и т. д.) и вода, используются многократно (обзор: Odum, 1986).
Ежегодно фотосинтезирующие организмы продуцируют до 1017 т (около 100 млрд. т) органического вещества. За это же время весьма значительное количество вещества разрушается и превращается в углекислый газ и воду в результате дыхания растений (Vallentyne, 1962). Предполагается, что преобладание органического синтеза над дыханием явилось главной причиной уменьшения содержания углекислого газа в атмосфере и накопления в ней кислорода, уровень которого в настоящее время достаточно высок. Именно это и способствовало эволюции и развитию высших форм жизни на Земле (обзор: Odum, 1986).
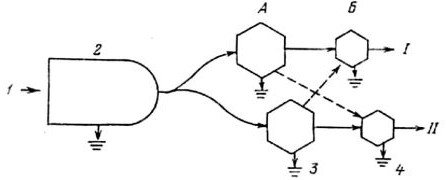
Рис. 1.6. Модель потока энергии, показывающая связь между пастбищной и детритной трофическим цепями (по: Odum, 1986).
I — пастбищная трофическая цепь; II — детритная трофическая цепь.
А — растительноядные, Б — хищные животные.
1 — солнечный свет; 2 — растения; 3 — потребители детрита; 4 — хищники.
Перенос энергии пищи от ее источника — аутотрофов (растений) через ряд организмов путем поедания одних организмов другими составляет трофическую цепь. При каждом очередном переносе до 80–90 % потенциальной энергии переходит в тепло. В связи с этим чем короче трофическая цепь, тем большее количество энергии доступно для популяции.
Формально трофические цепи можно разделить на пастбищную, начинающуюся от растений и идущую к растительноядным животным, и детритную, которая идет от мертвого органического вещества к микроорганизмам, а затем к детрофагам и хищникам (рис. 1.6). Важно, что трофические цепи не изолированы, а тесно переплетены, образуя так называемые трофические сети.
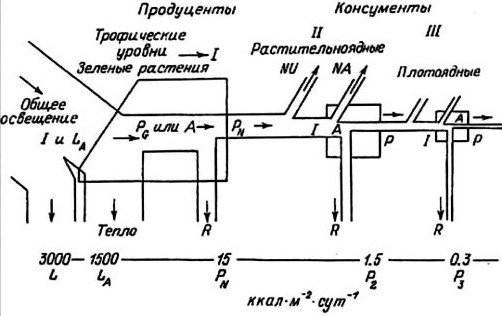
Рис. 1.7. Упрощенная схема потока энергии на трех трофических уровнях в линейной трофической цепи (по: Odum, 1986).
I, II, III — трофические уровни. I — общее поступление энергии; LA — свет, поглощаемый растительным покровом; PG — валовая первичная продукция; А — общая ассимиляция; PN — чистая первичная продукция; P — вторичная продукция консументов; NU — неиспользуемая (накапливаемая или экспортируемая) энергия; R — дыхание. Цифры внизу — порядок величины потерь энергии при каждом переносе, начиная с поступление солнечного излучение в количестве 3000 ккал. м-2?сут-1
Считается, что организмы, получающие энергию от Солнца через одно и то же количество этапов, принадлежат к одному уровню. При этом популяция может занимать не только один, а несколько уровней в зависимости от используемых источников энергии. Условно можно выделить 4 трофических уровня: 1) про дуценты (зеленые растения); 2) первичные консументы (травоядные); 3) вторичные консументы (первичные хищники, поедающие травоядных); 4) третичные консументы (вторичные хищники). Принято, что человек чаще всего одновременно является первичным и вторичным консументом, так как в его диету, как правило, входит пища как растительного, так и животного происхождения. В этом случае поток энергии разделяется между этими трофическими уровнями в соответствии с долями растительной и животной пищи. В превосходном обзоре Ю. Одума (Odum, 1986) представлена упрощенная схема потока энергии на трех уровнях (рис. 1.7). Как можно видеть, поток энергии через трофический уровень соответствует общей ассимиляции (А) на этом уровне, а последняя равна продукции (Р) биомассы плюс дыхание (R). На каждом следующем трофическом уровне поток энергии уменьшается. На первом трофическом уровне поглощается до 50 % падающего света, а в энергию пищи трансформируется только 1 % поглощенной энергии. На двух следующих уровнях вторичная продуктивность составляет лишь 10 % от предыдущей. В целом средняя эффективность переноса энергии между трофическими уровнями составляет 20 % и менее. Существует предположение, что регуляция первичной продукции зависит от числа звеньев в трофической цепи (Smith, 1969).
Что касается трофических цепей, то в большинстве случаев в каждом следующем звене поток доступной энергии уменьшается на один порядок. Равновесие между синтезом и деструкцией веществ — необходимое условие поддержания жизни в планетарном масштабе и существования каждого вида в отдельности.
Живые организмы, представляющие собой активную силу, действующую сегодня, составляют лишь сравнительно небольшую часть биосферы. Поэтому хотя носителями жизни являются отдельные организмы, жизнь, согласно В. И. Вернадскому (1926, 1965, 1980), возможна лишь как планетарное явление, как форма существования биосферы с обязательным для нее круговоротом веществ и потоков энергии, так называемым биотическим круговоротом. Этот круговорот определяется как метаболическими процессами, происходящими в отдельных организмах, так и связями между ними. Связи между различными организмами также в значительной мере являются трофическими.
Биотический круговорот в целом можно рассматривать как трофический процесс, а сами организмы включены в трофические цепи, точнее трофические сети, где каждый вид использует определенные источники питания и вместе с тем сам служит пищевым объектом. Источники питания, количество, свойства и доступность пищи в значительной мере лимитирует распределение и численность любой популяции, а также во многом определяют ее эволюционную судьбу. Следовательно, одним из условий устойчивости биосферы является биотический круговорот, или цикл, в который как важный компонент входят трофические взаимодействия. Последние обеспечивают образование биологической массы, ее миграцию и модификацию, а также постепенную деградацию до неорганических элементов, которые вновь включаются в начальные звенья цикла. Многие неорганические вещества превращаются в органические и трансформируются в метаболических звеньях трофических цепей, а затем в идеальных случаях возвращаются в метаболические звенья компонентов биотического круговорота. (Закономерности круговорота веществ и энергии в биосфере, проблема трофических связей и их роли в биосфере, ее эволюция и ряд других вопросов детально рассмотрены в ряде обобщающих сводок: Вернадский, 1926, 1965, 1980; Биосфера, 1972; Будыко, 1984; Уголев, 1985, 1986а, 1987а; Аксенов, 1986; Добровольский, 1986; Соколов, 1986; Allen, Nelson, 1986; Odum, 1986; Лапо, 1987; Green et al., 1990, и др.)
Таким образом, трофические связи являются обязательным элементом жизни как биосферного явления, а проблема происхождения жизни на Земле в конечном итоге сводится к вопросу об образовании первичной биосферы с ее первичными трофическими взаимоотношениями (см. гл. 9). Этот вопрос был поставлен еще В. И. Вернадским, как и вопрос о трофических связях в современной биосфере.
Понимание биосферы как трофосферы (Уголев, 1986а), состоящей из различных трофоценозов с их цепными и разветвленными связями, обеспечивающих циркуляцию веществ и энергии, позволяет решать проблемы охраны окружающей среды и поддерживать экологическое равновесие на основе анализа трофических соотношений и их сохранения. В некоторых случаях при нарушении трофических цепей возможно их восстановление за счет включения недостающих звеньев.
Следует обратить внимание на несколько аксиоматических положений: 1) в основе энергетики жизни и образования органических веществ лежат преимущественно процессы фотосинтеза, которые осуществляются за счет солнечной энергии; 2) основная часть энергии расходуется в результате метаболизма самими аутотрофными организмами; 3) лишь небольшая часть аккумулированного материала (примерно 10 %) переходит в следующее звено трофической цепи, и т. д. Следовательно, как уже отмечено выше, лишь небольшая часть энергии, накопленной в органических веществах, передается по трофическим цепям. Однако именно эта неметаболизированная часть и является самой важной, так как она определяет биологический состав биосферы, ее единство, гомеостаз и многие другие важнейшие свойства (обзор: Odum, 1986).
В классическом труде «Биосфера», опубликованном в 1926 г., В. И. Вернадский писал, что на земной поверхности нет химической силы, более постоянно действующей, а потому и более могущественной по своим конечным последствиям, чем живые организмы, взятые в целом. Следует отметить, что живые организмы необходимо рассматривать не как сумму автономных сил (в особенности трофических), а как системы, в которых активность организмов и популяций включена в определенные взаимодействующие и взаимосвязанные звенья.
Когда В. И. Вернадский создавал теорию биосферы, концепция трофических цепей базировалась на упрощенных и неполных представлениях, не позволяющих дать систематическое описание механизмов ассимиляции пищи, тем более в рамках единой науки — трофологии. Благодаря достижениям биологии последних лет представляется возможность охарактеризовать все процессы, происходящие в трофических цепях, на основании трех основных типов пищеварения, а также нескольких типов транспорта. Эти достижения позволяют также показать, что место вида в трофической цепи определяет его существование.
В сущности биология и физиология организмов тесно связаны с видовыми особенностями питания и добывания пищи. Экологи, и в частности Ю. Одум (Odum, 1986), подчеркивают, что главная побудительная причина активности всех животных — поиски нужной пищи в достаточном количестве. Твердо установлено, что структура и деятельность сообществ в значительной мере зависят от наличия пищи, а видовые и индивидуальные особенности многих процессов связаны с местом вида в трофической цепи и способом добывания пищи.
Парадоксальной на первый взгляд кажется другая связь: любой организм (точнее, вид) приспосабливается к тому, чтобы самому служить источником пищи (Уголев, 1980, 1986а). Для этого организм должен обладать такими свойствами, как определенная фагичность, т. е. доступность для другого организма в качестве источника пищи, а также трофичность, т. е. способность быть ассимилированным другим организмом. Некоторые рыбы, насекомые и растения, будучи вполне фагичными для многих организмов, из-за содержания в них токсических веществ не обладают трофичностью для ряда других живых существ.
На первый взгляд, такое положение трофологии может показаться и телеологичным, и противоречащим действительности, так как существует много данных в пользу того, что организмы вырабатывают специальные способы защиты от потенциального хищника.
Эта сторона проблемы детально освещена в прекрасном обзоре К. Шмидт-Ниельсена (Schmidt-Nielsen, 1982). Однако анализ показывает, что сочетание фагичности и трофичности обеспечивает процветание вида (разумеется, если оно не переходит определенных границ). Отсюда вытекают некоторые неожиданные следствия, в частности взаимная адаптация так называемых трофических партнеров (например, строгая взаимная зависимость свойств и численности популяций хищника и его жертвы). Так, в соответствии с гипотезой естественного равновесия (Hairston et al., 1960), выеданию биомассы травоядными животными препятствуют хищники, ограничивающие численность этих травоядных. Только в таком случае возможна определенная стабильность популяции жертвы за счет того, что хищники будут питаться преимущественно ее больными, дефектными и стареющими членами. Совершенствование же жертвы может привести хищника к гибели от голода и вследствие этого к ухудшению популяции жертвы из-за отсутствия контроля со стороны хищника. Анализ свойств этой пары демонстрирует важную роль источника пищи и наличие обратной связи. Это было продемонстрировано в невольных экспериментах человека, когда, например, в каком-либо регионе полностью уничтожалось поголовье хищника, а в другом происходила искусственная интродукция хищника с помощью специальных мероприятий по охране этих животных.
Взаимное приспособление в последовательных парах хищник-жертва можно проиллюстрировать некоторыми примерами, поражающими сложностью и точностью адаптаций. В частности, Т. Торнтон охарактеризовал способ, каким хищники оберегают вид, которым они питаются. Морские звезды и офиуры имеют период физиологического голодания в 1–2 месяца, приуроченный к моменту оседания личинок пластиножаберных моллюсков — одного из основных пищевых объектов названных хищников. Личинки очень малы, и популяцию моллюсков морские звезды и офиуры могли бы истребить чрезвычайно быстро, но этого не происходит, так как хищники «теряют аппетит». Когда же за время голодания хищников масса личинок увеличивается на 2–3 порядка, у звезд и офиур наступает пора питания. Так с помощью периодического голодания хищники сохраняют источник своего питания (по: Уголев, 1980).
Примером взаимных адаптаций может служить эволюция взаимоотношений паразит-хозяин, при которой происходят уменьшение вредных для хозяина последствий паразитизма или инфекционного воздействия и переход к нейтральным или часто симбионтным взаимоотношениям.
Таким образом, необходимым условием процветания вида является его положение в трофической цепи. Это положение обеспечивается эффективностью взаимодействий не только с предшествующими, но и с последующими членами цепи. Другими словами, существенную роль играет не только источник питания и его эффективное поглощение, но и поедаемость данного члена цепи. В сущности механизмы и закономерности трофических взаимодействий являются одной из самых важных проблем трофологии.
В последние десятилетия накопились серьезные основания для утверждения, что в биоценозах представители различных групп организмов, в том числе далеких таксонов и даже различных царств, взаимодействуют не только с помощью массообмена, но и с помощью химических посредников. Примеры таких взаимодействий представлены в ряде блестящих обзоров (Rice, 1978; Сравнительная физиология…, 1978; Schmidt-Nielsen, 1982; Odum, 1986, и др.).
Регуляторные взаимодействия организмов, так называемая аллелопатия (термин предложил Н. Моlisch в 1937 г.), на всех уровнях эволюционной лестницы, включая микроорганизмы, распространены очень широко. Таким образом, в первом приближении аллелопатия сводится к способности организмов определенных видов выделять физиологически активные вещества, действующие на организмы других видов. Классическим примером аллелопатии может служить образование антибиотиков и фитонцидов. Сейчас обнаружены многочисленные как токсические, так и стимулирующие вещества, вырабатываемые всеми живыми существами (см. также 9.7). Следовательно, состав любого биоценоза зависит не только от прямых трофических взаимодействий, но и от взаимодействий регуляторного характера.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
10.2. Современная проблема: человек и биосфера
10.2. Современная проблема: человек и биосфера Рост воздействия человека на биосферу непосредственно связан с ростом его численности. Разговоры о демографическом взрыве не просто красивые фразы. Увеличение числа людей на планете за последнее столетие носит именно
Биосфера и микроорганизмы
Биосфера и микроорганизмы Все пространство на земном шаре, населенное живыми организмами, мы называем биосферой. Биосфера охватывает верхнюю часть земной коры, воды рек, озер, морей, океанов и нижнюю часть атмосферы. В воде она достигает глубины 10 000 м. В почву дальше всех
Биосфера
Биосфера Биосфера представляет собой совокупность живых организмов Земли. Она охватывает нижнюю часть атмосферы, всю гидросферу и верхние слои литосферы. Живые организмы биосферы и их среда обитания образуют динамичную единую систему.Термин «биосфера» был введен
Глава 10. БИОСФЕРА
Глава 10. БИОСФЕРА 10.1. Понятие о биосфере Идея о влиянии жизни на природные процессы на огромных пространствах Земли была впервые научно обоснована на рубеже XIX и XX столетий в трудах В. В. Докучаева, который указал на зависимость типа почвообразования не только от климата,
Виды, биосфера и человек
Виды, биосфера и человек Книга Дэвида Эттенборо посвящена разнообразию форм жизни на Земле и путям возникновения этого разнообразия в процессе исторического развития жизни на нашей планете, то есть в процессе биологической эволюции.Удивительное разнообразие живого
3.4. Теория адекватного питания как составная часть трофологии
3.4. Теория адекватного питания как составная часть трофологии Достижения биологии и, в частности, ее физико-химических направлений служат основой для быстрого прогресса наших знаний в области физиологии и биохимии процессов ассимиляции пищи. Были сделаны крупнейшие
3.5. Биосфера как трофосфера
3.5. Биосфера как трофосфера Жизнь на Земле возможна лишь как планетарное явление, как форма существования биосферы с обязательным для нее кругооборотом веществ и потоков энергии — биотическим круговоротом. Равновесие между синтезом и деструкцией веществ — необходимое
28. Биосфера – глобальная экосистема
28. Биосфера – глобальная экосистема Вспомните!Какие уровни организации живой природы вам известны?Что такое биосфера?Каковы её границы?Многочисленные экосистемы нашей планеты не изолированы друг от друга. Даже между очень разными сообществами происходит постоянный
30. Биосфера и человек
30. Биосфера и человек Вспомните!Как протекала эволюция биосферы?Какова роль человека в биосфере?Ранние этапы развития человечества. Влияние человечества на биосферу началось в тот момент, когда люди перешли от собирательства к охоте и земледелию. По мнению учёных, уже в
31. Основные экологические проблемы современности
31. Основные экологические проблемы современности Вспомните!Какие глобальные экологические проблемы вам известны?Приведите примеры экологических проблем вашего региона.Загрязнение атмосферы. Одной из самых острых экологических проблем в настоящее время является
1.2. Предмет и задачи трофологии
1.2. Предмет и задачи трофологии Предметом трофологии являются общие закономерности ассимиляции жизненно необходимых пищевых веществ на всех уровнях организации биологических систем — от уровня клетки, органа, организма до популяций, биоценозов и биосферы в целом.
1.5. Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера
1.5. Популяционные, экологические и эволюционные проблемы трофологии. Биосфера как трофосфера С деятельностью живых систем связана та часть поверхности Земли, которая объединена под названием биосферы. Биосфера, являющаяся самой крупной экосистемой, представляет собой
Глава 9. Трофические аспекты происхождения жизни в свете трофологии. Некоторые биологические аспекты трофологии
Глава 9. Трофические аспекты происхождения жизни в свете трофологии. Некоторые биологические аспекты трофологии 9.1. Вводные замечания Из предшествующего изложения ясно, что трофология в целом останется собранием прикладных направлений, если не будут учтены многие
1.2. Предмет и задачи трофологии
1.2. Предмет и задачи трофологии Предметом трофологии являются общие закономерности ассимиляции жизненно необходимых пищевых веществ на всех уровнях организации биологических систем — от уровня клетки, органа, организма до популяций, биоценозов и биосферы в целом.
Глава 9. Трофические аспекты происхождения жизни в свете трофологии. Некоторые биологические аспекты трофологии
Глава 9. Трофические аспекты происхождения жизни в свете трофологии. Некоторые биологические аспекты трофологии 9.1. Вводные замечания Из предшествующего изложения ясно, что трофология в целом останется собранием прикладных направлений, если не будут учтены многие
Вода и биосфера
Вода и биосфера Внешняя оболочка Земли занята биосферой. И вполне правильно, когда биосферу называют еще «областью жизни» или «живым покровом» Земли. Это огромное пространство, включающее атмосферу, гидросферу и литосферу, населяют различные виды живых организмов.